
在维管植物中,根内皮层作为皮层最内侧的细胞层,包裹维管组织并发挥关键的过滤功能。这一功能依赖于内皮层细胞壁局部木质素沉积形成的凯氏带,迫使营养物质通过主动运输跨膜进入维管组织,从而确保植物对养分吸收的选择性和调控能力。凯氏带的形成始于根发育早期,与根毛伸长同步。在豆科植物中,凯氏带形成区域与根瘤菌易感区空间上重合,但二者之间的内在联系尚不明确。此外,根瘤维管组织与主根维管组织相连,是植物与根瘤菌进行氮碳营养交换的重要通道。类似地,根瘤维管组织也被含有凯氏带的内皮层包裹,但其具体生理功能此前尚未阐明。
百脉根凯氏带突变体揭示其在根瘤形成中的关键作用
德国马克斯普朗克植物育种研究所Tonni Grube Andersen团队联合多国科研人员在《Science》发表研究论文,题为“Apoplastic barriers are essential for nodule formation and nitrogen fixation in Lotus japonicus”。研究通过鉴定百脉根(Lotus japonicus)凯氏带相关突变体,揭示了凯氏带不仅参与调控根瘤形成,还在维持根瘤固氮功能中发挥重要作用。
前期研究表明,拟南芥中凯氏带的形成受MYB36转录因子、SCHENGEN3(SGN3)受体激酶及其下游SGN1协同调控。在百脉根中,Ljmyb36、Ljsgn1和Ljsgn3突变体均缺失凯氏带,表明该调控通路在百脉根中具有保守性(图1)。
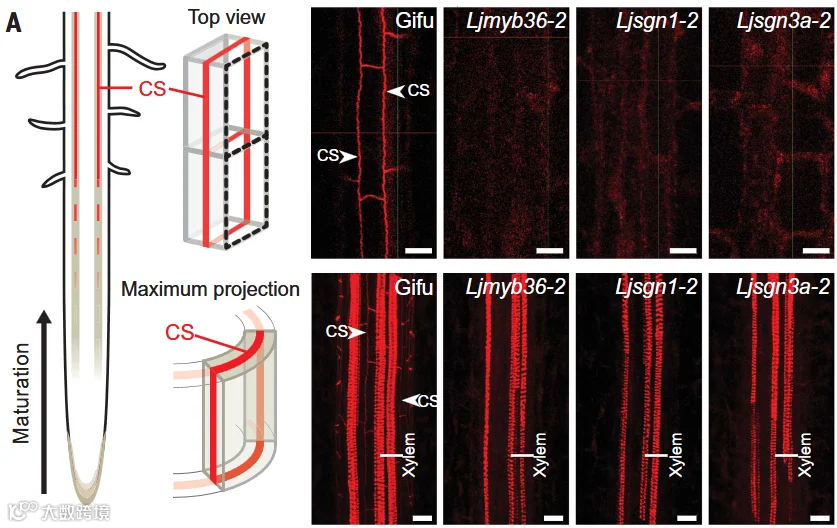
图1 与野生型Gifu相比,Ljmyb36、Ljsgn1和Ljsgn3突变体表现出凯氏带缺失表型。红色信号代表碱性品红染色后的木质素,凯氏带(CS)呈现网格状结构。维管组织的木质部因含木质素而被染色。
凯氏带通过系统性信号调控根瘤形成
研究发现,上述突变体存在结瘤延迟、结瘤数量减少及固氮能力下降等共生缺陷,根源在于结瘤自调控系统(Autoregulation of Nodulation)紊乱。在低氮条件下,凯氏带可诱导根部表达氮饥饿信号小肽CEP1,促进地上部分合成microRNA2111(miR2111),进而抑制其靶基因——结瘤负调控因子TOO MUCH LOVE(TML)在根瘤菌易感区的表达,从而促进根瘤发生(图2)。该机制揭示了凯氏带除调控养分吸收外,还参与植物低氮响应及根瘤发生的系统性调控。
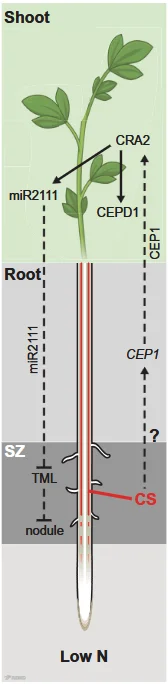
图2 凯氏带系统性调控根瘤形成的模型。CS:凯氏带;SZ:根瘤菌易感区。
根瘤凯氏带维持氮碳高效交换
根瘤菌将氮气固定为氨,植物将其转化为天门冬酰胺等氨基酸,并通过根瘤维管组织向外运输,提供氮源;同时,植物向根瘤输送蔗糖等碳源以支持固氮活动。研究证实,根瘤凯氏带的形成机制与根部一致,且在Ljmyb36突变体中,根瘤内天门冬酰胺、蔗糖及营养元素的分布显著异常(图3),表明凯氏带通过调控根瘤内部物质运输与分配,保障氮碳高效交换,维持正常固氮功能。
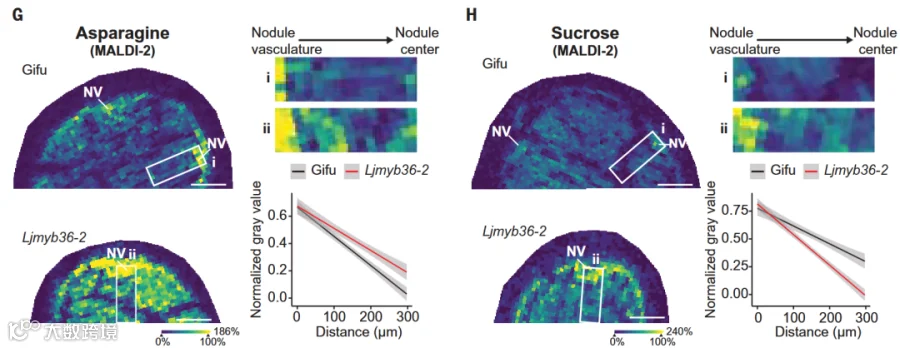
图3 与野生型Gifu相比,根瘤凯氏带缺失突变体Ljmyb36中天门冬酰胺和蔗糖的分布发生显著改变。NV:根瘤维管组织。
该研究确立了凯氏带作为研究植物-微生物共生关系中空间屏障功能的理想模型系统,为深入理解植物与微生物互作过程中物理屏障的生理意义提供了新的理论框架。丹麦哥本哈根大学、德国科隆大学和慕尼黑大学的研究人员亦对本工作作出重要贡献。论文第一作者为申德峰博士,通讯作者为Tonni Grube Andersen博士。
原文地址:https://www.science.org/doi/10.1126/science.ado8680