

论文概览
小麦氮素利用效率研究进展
戎洛坤1,2,赵荣斌3,林平4,王磊1*
1 中国科学院遗传与发育生物学研究所 农业资源研究中心
2 中国科学院大学
3 石家庄市栾城区农业农村局
4 菏泽市农业科学院
摘要:氮素利用效率(nitrogen use efficiency, NUE)是小麦高产与资源可持续利用的关键指标。本文总结了近年来小麦氮素利用效率的研究进展,聚焦基因型差异、生理机制、分子调控及栽培管理措施,分析了氮素吸收、转运、同化与再分配的关键调控节点,并总结了当前研究的瓶颈与未来方向。结合新型生物技术手段与精准农艺管理,提出协同提升小麦产量与氮素利用效率的策略,为氮高效品种选育及绿色生产提供理论支持。
氮素利用效率(nitrogen use efficiency, NUE)是衡量作物对氮肥吸收、利用和转化能力的重要指标,对于提高小麦产量、优化氮肥管理、降低环境污染具有重要意义。全球小麦生产面临氮肥利用率低、施氮过量导致的资源浪费与环境污染等问题,提高NUE成为实现可持续农业的关键途径。近年来,围绕小麦NUE的研究在基因型差异、生理机制、分子调控及栽培管理等方面取得了重要进展。基因型筛选方面,不同品种在氮素吸收效率(nitrogen uptake efficiency, NUpE)、氮素生理利用效率(nitrogen utilization efficiency, NUtE)及氮收获指数(nitrogen harvest index, NHI)上表现出显著差异,特定基因位点的挖掘和功能验证为氮高效品种的培育提供了理论依据。在生理机制层面,氮素(nitrogen, N)的吸收、转运、同化与再分配涉及一系列酶活性调控与代谢途径优化,根系构型、激素信号、光合能力在调控NUE方面发挥核心作用。分子机理研究完善了氮素调控网络,为小麦氮素高效利用的遗传改良提供了新方向。同时,精准农业技术的发展,如无人机多光谱遥感、土壤氮传感器、变量施肥系统及缓控释肥的优化应用,提高了氮肥管理的精准性,减少了氮素流失(图1)。本综述聚焦近几年小麦NUE研究的最新进展,从基因型差异、生理代谢、分子调控及栽培管理这4个方面总结现有研究成果,分析当前存在的瓶颈,并探讨未来提高NUE的研究方向,以期为小麦氮高效育种及绿色生产提供理论依据。
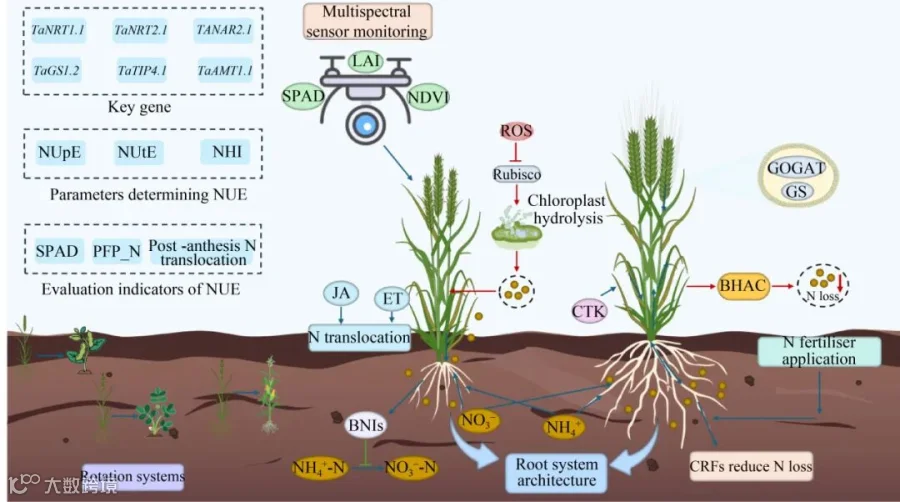
图1 小麦氮素利用效率综合调控模式图 关键基因通过调控氮素的吸收、转运及同化过程提升NUE;激素信号通路进一步优化氮素再分配。精准施氮技术结合智能检测手段,实现氮肥的高效利用;轮作系统通过生态调控机制减少氮素损失。N:氮;NUE:氮素利用效率;NUpE:氮素吸收效率;NUtE:氮素生理利用效率;NHI:氮收获指数;SPAD:旗叶叶绿素相对含量;PFP_N:氮肥偏生产力;BNIs:生物硝化抑制剂;GS:谷氨酰胺合成酶;GOGAT:谷氨酸合酶;ROS:活性氧;ET:乙烯;JA:茉莉酸;CTK:细胞分裂素;BHAC:β-羟基天冬氨酸循环;LAI:冠层叶面积指数;NDVI:归一化植被指数;CRFs:缓控释肥。
Figure 1 Comprehensive regulation of wheat NUE. Key genes enhance NUE by regulating nitrogen uptake, transport, and assimilation processes. The hormone signaling pathway optimizes nitrogen redistribution. Precision nitrogen management integrated with intelligent detection techniques enables the efficient utilization of nitrogen fertilizer. The rotation system reduces nitrogen loss via ecological regulation mechanisms. N: Nitrogen; NUE: Nitrogen use efficiency; NUpE: Nitrogen uptake efficiency; NUtE: Nitrogen utilization efficiency; NHI: Nitrogen harvest index; SPAD: Soil and plant analyzer development; PFP_N: Partial factor productivity of nitrogen fertilizer; BNIs: Biological nitrification inhibitors; GS: Glutamine synthetase; GOGAT: Glutamate synthase; ROS: Reactive oxygen species; ET: Ethylene; JA: Jasmonic acid; CTK: Cytokinin; BHAC: β-hydroxyaspartate cycle; LAI: Leaf area index; NDVI: Normalized difference vegetation index; CRFs: Controlled-release fertilizers.
1 氮素利用效率的含义与评价指标
NUE是指单位可用氮素对作物产量的比值,通常通过氮输出与氮输入之间的比值来衡量。NUE的计算公式为:NUE=N输出量/N输入量,其中N输出量是指作物收获中的氮含量,N输入量包括肥料投入、大气沉降和土壤中矿化产生的氮素[1-2]。NUE包括作物对氮素的吸收、利用和再分配过程,其受氮素吸收效率(NUpE)、氮素生理利用效率(NUtE)和氮收获指数(NHI)等参数影响[3]。在NUE的测定和评价过程中,开花后氮素转运量、氮肥偏生产力(partial factor productivity of nitrogen fertilizer, PFP_N)和旗叶叶绿素相对含量(soil and plant analyzer development, SPAD)动态变化等多个指标被广泛应用于不同作物。
1.1 氮素吸收效率(NUpE)
氮素吸收效率是植物从氮源吸收的氮总量,其计算公式为:NUpE=地上部氮积累量/土壤供氮量。NUpE反映作物根系从土壤中获取氮素的能力,取决于根系生长状况、土壤氮素供应水平及水肥管理条件[4-5]。根系生长旺盛、分布深广的作物能够从更大范围内吸收氮素以提高NUpE。土壤氮素形态也会影响氮素吸收,铵态氮和硝态氮的供需平衡直接决定氮素进入根系的效率[6]。
1.2 氮素生理利用效率(NUtE)
氮素生理利用效率的定义为单位氮素积累量对应的籽粒产量,其计算公式为:NUtE=籽粒产量/植株总氮积累量。NUtE是评价作物将吸收的氮素用于生物量积累和籽粒形成能力的指标,受作物光合作用、碳氮代谢及氮素在植株内部再分配的影响[7-8]。小麦高效品种NUtE可达50–60 kg/kg N,低效品种仅30–40 kg/kg N,高NUtE品种能够在较低氮素水平下维持较高的干物质生产能力。NUtE反映单位氮素积累量对产量的贡献,与品种的耐低氮能力密切相 关[9]。高效作物品种在低氮条件下仍能保持较高NUtE,提高氮素利用效益[10]。NUtE能够表征氮同化与碳代谢的协同效率,用于不同品种耐低氮能力评估。
1.3 氮收获指数(NHI)
氮收获指数是指籽粒氮含量占植株总氮含量的百分比。NHI代表了植株内氮素向可收获部分的分配比例,反映氮素转运至籽粒或经济产量部分的效率[11]。高产小麦品种NHI明显高于低效品种,较高的NHI意味着更多氮素从叶片和茎秆向籽粒转移,最终将提高作物产量[12-13]。NHI能够评价氮素向籽粒的分配效率,从而可以将其作为筛选高产育种的重要指标之一。
1.4 开花后氮素转运量
开花后氮素转运量是指开花后茎叶氮素向籽粒的转运量,其计算公式为:转运量=开花期茎叶氮量‒成熟期茎叶氮量。开花后氮素转运量能够评估氮素再分配能力,适用于测定生殖生长期氮素从营养器官向籽粒的转移量,其与籽粒蛋白质含量密切相关。研究表明,氮高效品种转运量能达到60 kg/hm2以上,占籽粒氮总量的70%–80%[14],开花后氮素转运量能够直接影响作物品质。
1.5 氮肥偏生产力(PFP_N)
氮肥偏生产力是指单位施氮量对应的籽粒产量,其计算公式为:PFP_N=籽粒产量/施氮量。PFP_N是衡量单位施氮量对应的籽粒产量及评价田间氮肥效率的关键指标[15]。优化施肥条件下小麦PFP_N能够达到40–50 kg/kg,传统施肥PFP_N仅有25–30 kg/kg[16]。较高的PFP_N表明作物能在较少氮肥投入下获得较高产量,对田间氮肥管理具有指导意义,适用于精准农业技术评估。
1.6 旗叶叶绿素相对含量(SPAD)动态变化
旗叶叶绿素相对含量在灌浆期的动态变化一方面反映叶片氮素滞留能力与光合功能期的协同关系,另一方面反映作物氮素供应的时序特征[12]。氮高效品种灌浆期SPAD值下降速率明显小于氮低效品种[17],这种差异直接体现在光合功能持续性上,SPAD值下降速率较慢的品种能在灌浆期维持较长的光合功能期。
2 小麦氮素利用效率的遗传生理机制
2.1 氮素吸收与根系构型
氮高效品种在不同氮素水平下表现出显著的产量和生理差异,低氮条件下氮高效品种的产量损失远低于氮低效品种,这种优势源于根系结构、氮素转运能力和代谢酶活性的差异。根系分布和总根表面积等作物根系形态特征影响了NUpE,抗旱型小麦在深层土壤的根系分布较多,在低氮环境下依然能够维持较高的氮吸收水平;深根系品种在低氮环境下的优势明显,根系的垂直分布影响了NHI[18]。基因型差异在不同土壤氮素供应水平下表现出显著性,深层根系发育良好的品种能够更有效地吸收土壤中残留的硝态氮[19]。研究表明,根系生长的动态变化与氮素供应水平之间存在密切联系,根系形态的表型组学分析有助于筛选氮高效利用基因型[14]。根系构型决定了作物的氮素捕获能力,深根型品种在低氮和干旱条件下表现出更强的适应性[20]。作为冬小麦的‘济麦22’其根系构型优化表现为深层根系密度增加,该根系构型使20–60 cm土层的NUpE显著提高。深根系在水分受限环境下的作用尤为突出,能够持续获取深层土壤的残留氮素,提高了NUE[21]。根系构型优化的目标是形成兼具高氮吸收能力和低养分消耗的模式,使作物在不同土壤环境中均能保持较高的NUE。
氮素吸收和转运的遗传调控涉及NRT1.1、NRT2.1、NLP7等多个转录调控因子[22-26],这些转录因子在调控作物NUE的过程中发挥了重要作用。高亲和力硝酸盐转运蛋白(nitrate transporter 2, NRT2)家族基因在氮信号转导过程中扮演关键角色,特定NRT基因的过表达增强了作物对氮素的响应能力[27]。Shi等[28]研究发现,在氮高效品种中,NRT基因的表达水平的提高增强了根系对外源氮素的敏感性,结合转录组数据分析,NPF、NRT2、NAR2等与氮代谢相关的基因,在不同氮素水平下表现出时序表达差异,基因调控网络的构建有助于理解氮素高效利用的分子机制。TaNRT1.1能够控制硝态氮的转运能力,基因突变可导致氮吸收效率的差异[22]。王沙沙等[29]解析了小麦硝酸盐转运蛋白基因TaNRT1.1的生物学功能,通过同源克隆的方法得到TaNRT1.1的同源基因,研究发现同源基因在小麦不同组织以及NUE不同的小麦品种中的相对表达量有明显差异。
硝态氮和铵态氮的吸收由不同基因控制,而且铵态氮与硝态氮的不同配比会影响根系吸收的选择性。高亲和力硝酸盐转运蛋白TaNRT2.1能够调控小麦的硝态氮吸收,这一过程需伴侣蛋白TaNAR2.1协助完成跨膜转运[11]。TaNRT2.1的磷酸化调控影响其与NAR2.1的结合强度,决定了NUpE。基因过表达研究表明,在高氮供应下,TaNRT2.1可显著提升氮素吸收能力,而在低氮条件下效果有限,表明该基因的作用依赖于环境氮浓度[23]。硝态氮和铵态氮的协同作用受植物激素调控,脱落酸(abscisic acid, ABA)信号可通过TaBG1水解ABA β-d-葡萄糖苷(ABA glucosyl ester, ABA-GE),激活TaNRT2.1和TaAMT1.1的表达,使小麦根系NUpE大幅度提高[30]。
根系的氮素捕获策略随环境变化而动态调整,氮素形态的分布驱动根系构型与吸收能力的优化[31]。在干旱胁迫下,轻度水分亏缺可促进小麦中下层根系延伸,提高深层氮素的吸收能力,而重度干旱则抑制整体根长,降低氮素获取能力。部分基因型能够通过根系分泌生物硝化抑制剂调节土壤氮素转化过程,延缓铵态氮向硝态氮的转化,减少硝态氮的流失[32]。氮高效基因型结合生物硝化抑制剂,可在土壤保持较高铵态氮水平的情况下提高铵氮的NUpE,进一步优化氮素利用策略[31]。
2.2 氮素转运同化与再分配
作物的氮素高效利用依赖于代谢途径、激素信号和环境因子等的综合调控以确保氮素从吸收到转运、再分配的全过程协调,最终影响籽粒蛋白质含量和产量。植物根系吸收的不同形式的氮源,还需要经过转运同化与再分配,才能被植物所利用。
韧皮部是氮素长距离运输的主要组织,其决定了籽粒氮积累的效率[33]。谷氨酰胺合成酶(glutamine synthetase, GS)在氮素同化和再转运过程中发挥核心作用,其活性水平直接影响籽粒蛋白质含量[34]。研究表明,GS活性与籽粒氮积累量呈正相关,灌浆期韧皮部液中的谷氨酰胺浓度决定了氮素向籽粒的输入速率[35]。TaGS1.2基因的过表达可显著提升籽粒蛋白质含量,转基因小麦品系中TaGS1.2表达量提高,使籽粒蛋白质含量增加,NHI提高[36]。GS1在非光合组织中优势表达,使其在叶片衰老期催化释放的铵离子转化为谷氨酰胺,促进氮素向籽粒的再分配,确保低氮环境下氮素供应稳定[33]。
不同生育期的氮素吸收特征受基因表达水平调控,氮素转运同化与再分配基因的时序表达影响了最终产量和品质。氮素转运受氮浓度、光照和激素信号的协同调控。籽粒蛋白质含量与氮素转运效率呈正相关,高蛋白品种在开花后吸收的氮素对最终籽粒蛋白质含量的形成具有显著贡献,其贡献程度大于其他生育阶段的氮素积累作用[37]。TaAMY3过表达增强α-淀粉酶活性,加速淀粉降解,为氮素同化提供碳源,使籽粒灌浆期NUE提升[38]。氮素供应不足时,籽粒灌浆速率降低,使氮利用率下降;而充足的碳氮供应可使籽粒灌浆速率提高,使籽粒蛋白含量增加。
而花后氮素再分配决定了籽粒品质,衰老叶片氮素释放量可占籽粒氮积累的60%以上[34]。液泡膜转运蛋白在氮素再分配过程中起关键作用,液泡作为氮素储存库,通过TaTIP4.1介导的NH4+外排将氮素释放至细胞质,再经谷氨酰胺合成酶/谷氨酸合酶(glutamate synthase, GOGAT)途径转化为谷氨酰胺,通过韧皮部运输至籽粒[39]。TaTIP4.1的过表达提高液泡NH4+外排速率,转基因小麦中TaTIP4.1表达量上调,使衰老叶片的NUE提升,籽粒氮积累量增加[40]。
衰老相关信号促进氮素再分配,活性氧(reactive oxygen species, ROS)积累触发Rubisco降解,加速叶绿体蛋白水解,释放氮素至韧皮部[41-42]。CND41蛋白酶在此过程中发挥作用,使氮素释放速率提高[43]。光合同化过程中,氮代谢相关酶的活性水平决定了氮素的最终分配,酰胺合成途径的调节直接影响作物的NUE。研究发现GS和GOGAT这类氮代谢关键酶在高效品种中表现出更高的活性水平,同时促进了氮素的同化与再分配[44]。灌浆期氮素同化能力差异导致品种间的产量表现出现分化,强筋小麦品种在灌浆期的氮素转运效率较高,在酰胺态氮供应条件下,旗叶GS活性显著增强,表现出更高的NUE,最终籽粒蛋白质含量提升显著[45-46]。
氮素代谢相关基因的调控机制涉及多层次的信号传导,激素信号在NUE中的作用逐步得到验证。乙烯(ethylene, ET)和茉莉酸(jasmonic acid, JA)可上调液泡膜转运蛋白基因的表达,增强氮素向籽粒的转运能力。细胞分裂素(cytokinin, CTK)信号延缓叶片衰老,影响氮素释放时机,使氮素的供应与籽粒灌浆进程同步。外源CTK处理可使衰老叶片氮素滞留时间延长,使籽粒蛋白质含量提高,确保氮素的高效利用[47]。
环境条件对氮素再分配产生影响,干旱和高温胁迫下,液泡膜H+-ATPase活性增强,维持跨膜质子梯度,保障氮素转运效率。低氮胁迫下,高效基因型的NUE依然保持稳定,而低效基因型的NUE则表现出显著下降。不同品种在土壤氮素供应充足或缺乏条件下的表现对比表明,氮素代谢的遗传调控决定了作物的生长适应性[28]。
2.3 碳氮代谢协同
氮素高效转运依赖于碳氮代谢的协同作用,NUtE反映作物单位氮素积累所能形成的产量水平,不同品种NUtE存在差异的本质在于碳氮代谢协同能力的强弱。高效品种通过碳氮代谢的动态平衡,在低氮条件下仍能维持叶片光合产物向籽粒的分配效率,并延缓叶片衰老,从而确保灌浆期氮素供应与碳同化的高效耦合[48]。作物碳氮代谢协同的核心在于Rubisco酶活性与氮素分配的动态平衡。近年来,通过遗传改良提升Rubisco羧化效率成为研究热点,Nazari等[42]将甘蔗RbcS基因导入水稻后,Rubisco羧化速率提高,从而使田间产量增加,在小麦中尚未有相关的遗传改良研究报道。另一方面,过量施氮会削弱Rubisco的CO2亲和力,导致光呼吸增强。优化氮素分配可缓解这一矛盾,适当调整氮肥施用时期,将冬小麦氮肥施用后移至拔节期,可使旗叶Rubisco活性提高,从而提高作物NUE[41]。
随着光呼吸代谢改造技术取得突破性进展,细胞质通路以及光呼吸蛋白之间形成复合物通过重新固定光呼吸释放的NH3,使水稻在低氮条件下的产量提高[49]。光呼吸旁路改造后的作物表现出更高的NUE,可以使光呼吸释放的CO2重新进入氨基酸合成途径,在低氮条件下的NUE和产量都有所提升[50]。Song等[51]研究发现,为避免光呼吸导致的碳损失,β-羟基天冬氨酸循环(β-hydroxyaspartate cycle, BHAC)以能量和氮的形式进行高效回收,从而将光呼吸副产物转化为C4代谢前体草酰乙酸,提高碳固定效率,减少氮素的损耗。氮素形态调控对碳氮平衡具有显著影响,增加硝态氮比例可缓解强光胁迫,减少甘氨酸脱羧酶造成的氮损耗,当叶片C/N比在一定范围内降低时,光呼吸氮素浪费大幅度减少[42,52]。水稻通过MAPK2调控GLO1磷酸化修饰,成功降低光呼吸ATP消耗,使光合效率提升[53]。不同作物在不同氮素管理模式下展现出不同的光氮耦合特征,合理优化碳氮代谢通路可减少氮素损耗,增强光合作物对氮肥的响应,使单位氮素投入产量最大化。
3 提高氮素利用效率的栽培管理措施
3.1 精准施氮及智能检测技术
基因型与施肥管理的协同优化是提高作物NUE的重要手段,精准施肥也能够提高NUE[54]。王文军等[55]研究发现,在砂姜黑土区小麦-玉米轮作制中,不同有机肥替代化肥的比例会影响小麦产量,在一定范围内,有机肥替代比例与小麦产量呈正相关。当在氮肥中加入脲酶抑制剂或硝化抑制剂时,一方面可有效地减缓氮肥酶解过程,同时延长氮肥的扩散时间,从而提高氮肥的有效利用率[32,56]。另一方面,侧深施肥技术、液体氮肥精准喷施结合GPS导航技术等,也显著提升了氮肥利用率[57-58]。
冠层光谱信息可以反映作物在不同生育阶段的氮素动态变化,不同品种在光谱曲线上展现出差异化特征[59]。小麦品种在灌浆期的冠层光谱特征与NUpE显著相关,冠层叶面积指数(leaf area index, LAI)和归一化植被指数(normalized difference vegetation index, NDVI)在拔节期与NUpE的关系尤为明显[60]。氮高效品种在光合时期的红边波段反射率较低且稳定性更高,表明氮高效品种叶片对氮素的保持能力更强[61-62]。光谱分析结合SPAD测定能够准确捕捉氮素的动态变化,研究表明,在不同氮素供应水平下,氮高效基因型在光合作用关键时期的SPAD值下降速率较慢,确保了生殖生长期氮素的持续供应,使NUE得到提升[17,63]。
无人机遥感与土壤氮素传感技术结合的变量施肥系统可以动态监测作物对氮的需求,在合适时间施加氮肥,从而提高NUE[58]。另外,无人机遥感结合多时相影像分析及机器学习模型,能够精准捕捉冠层光谱特征信息的变化,有效识别氮高效表型,同时也能精准预测NUE,显著提升氮高效品种的筛选效率[64-66]。
3.2 农艺措施优化
小麦-豆科轮作系统通过根瘤菌固氮作用显著降低对化学氮肥的依赖[67],研究表明,小麦-箭筈豌豆轮作系统可固氮50–200 kg/hm2,满足豆科作物大部分的氮素需求,同时使小麦氮素利用率提升。在优化根系系统配置方面,小麦-玉米轮作系统通过根系分层利用氮素(玉米吸收深层残留氮;小麦利用表层氮),从而整体提高NUE[68]。同样,小麦-苜蓿轮作系统则利用苜蓿深根系统减少深层氮淋失,使后续小麦季氮吸收量增加[41]。
长期轮作对土壤的氮循环具有持续调控作用。长期单施化肥会降低土壤固氮能力,而结合有机肥管理,既可以增加有机氮的含量,优化氮素供应,又能恢复土壤微生物功能,使固氮功能基因丰度提升,从而提高NUE[69]。
4 小麦氮素利用效率研究的瓶颈
小麦NUE的研究面临多方面瓶颈。在氮素转化与土壤供氮调控方面,虽然采用同位素示踪技术揭示了铵态氮与硝态氮的转化规律,但长期过量施氮导致的土壤酸化问题仍缺乏普适性解决方案;精准监测技术中无人机遥感模型在不同品种、环境及生长阶段的泛化能力仍待提升,需依赖大量样本训练来进行优化;禾本科-豆科轮作的农艺措施虽能减少氮肥投入,但对后茬作物氮吸收的长期影响仍需验证;生理机制上,碳氮代谢协同调控的复杂性及品种间氮素分配差异显著,导致氮肥施用缺乏统一标准。总体而言,技术适应性不足、模型泛化能力有限、农艺措施与生态平衡的矛盾及生理 机制复杂性都制约了小麦NUE研究的突破性进展。
5 结语
小麦NUE受基因型、生理代谢及施肥管理的影响。高NUE品种在根系结构、氮素吸收能力及光合氮代谢调控方面表现出明显优势。关键基因如TaNRT1.1、TaGS1.2、TaAMT1.1、TaTIP4.1等在不同生育阶段调控氮素的吸收、转运、同化或代谢,调控植株对氮素的利用能力。近年来,基因组学、转录组学以及表型组学等多组学的联合分析推动了氮高效基因的筛选与育种进程,基因编辑技术的应用进一步加速了小麦NUE机制的完善。同时,光合-氮素代谢的耦合、氮素同化酶活性的提升、根际微生物的调控对提高NUE具有重要影响。另外,精准农业技术的发展,使变量施肥、缓控释肥及增效剂的应用优化了传统的氮素供应模式,提高了作物的NUE。

更多内容,详见最下方【阅读原文】
推荐阅读
当年目次

2025年41卷第1期

2025年41卷第2期

2025年41卷第3期
2025年41卷第4期

2025年41卷第5期

2025年41卷第6期

2025年41卷第7期
2025年41卷第8期

2025年41卷第9期

2025年41卷第10期
欢迎加入科微学者交流群



扫描二维码



期刊订阅
1.直接联系联合编辑部订购(订购全年期刊可享9折优惠,当期作者购刊可享5折优惠),发行部 E-mail:bjb@im.ac.cn ;Tel:010-64807336;
2.各地邮局订阅:邮发代号82-13;
3.网上购买:搜淘宝店、微店店铺名称:中科期刊(订阅及销售过刊);科学出版社期刊发行部:联系电话010-64017032 64017539;
或扫描下方二维码:


——中文期刊联合编辑部出品