责编 | 兮
细菌的防御系统包括限制-修饰系统、CRISPR系统、感染流产系统等,是细菌抵御天然宿敌噬菌体的重要防御性武器。早在上个世纪,科学家们就对限制-修饰系统展开了研究,由此开发的限制性内切酶广泛地应用于分子生物学领域。DNA磷硫酰化修饰是一类全新的限制-修饰系统,与传统的DNA甲基化限制-修饰系统相比,该系统用硫原子替代了磷酸骨架上非桥连的氧原子。目前已知两种磷硫酰化系统(细菌与古菌)的修饰部分均为Dnd蛋白编码,仅在限制部分不同。
近日,武汉大学药学院王连荣团队在Nature Microbiology上发表了题为SspABCD-SspE is a phosphorothioation-sensing bacterial defence system with broad anti-phage activities的研究论文,报道了SspABCD-SspE组成的一种全新的磷硫酰化限制-修饰系统并对其防御机理进行了深入探究。
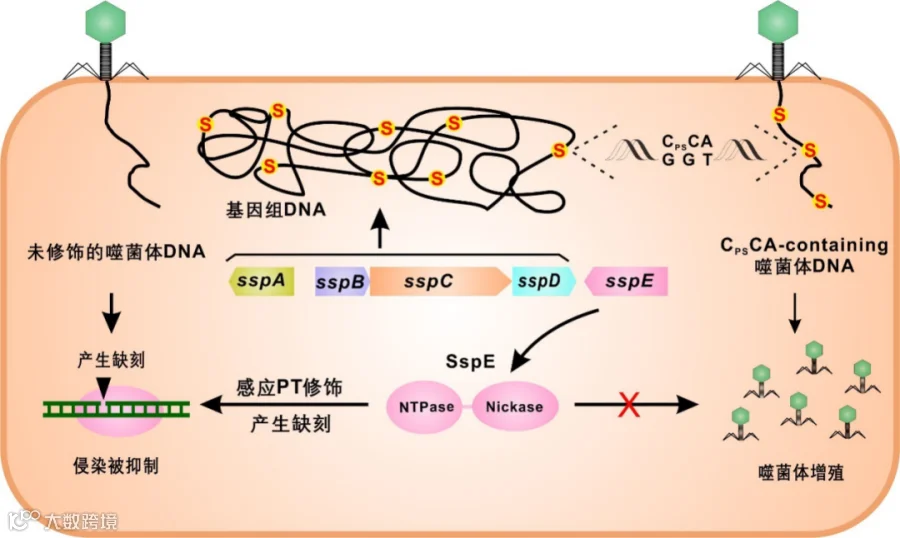
图1. DNA单链磷硫酰化修饰-感应修饰限制系统的工作机理
以往发现的磷硫酰化修饰均位于基因组双链DNA上。海洋弧菌FF75具有一类特殊的高频单链磷硫酰化修饰并位于CPSCA核心序列。然而,在 FF75的基因组上没有找到dnd基因簇,这促使研究者推断FF75中可能存在一套全新的编码单链磷硫酰化修饰的基因簇。通过比较基因组分析,对一段疑似基因簇进行了基因敲除,随后将这个基因簇的四个基因异源表达到大肠杆菌DH10B中重现了单链磷硫酰化CPSCA修饰,证明这四个基因正是单链修饰编码基因。研究者以单链修饰(Single-stranded phosphorothioation)来命名四个基因为sspA、sspB、sspC与sspD。
鉴定出修饰基因后,研究者尝试探索该修饰的生理意义。实验过程中一个意外发现引起了他们的注意:4摄氏度保存的养菌平板,仅野生型FF75菌株可以缓慢生长,磷硫酰化修饰丧失的突变株生长受到抑制。转录组结果显示这些生长抑制的突变株中SOS基因转录水平普遍上升,暗示在FF75中很可能存在一套识别CPSCA修饰的限制系统。经过蛋白组比对及大范围基因敲除,鉴定出在sspBCD下游的sspE基因编码了单链磷硫酰化的限制系统。将单链磷硫酰化限制-修饰基因簇异源表达到大肠杆菌中,可以赋予宿主抵抗包括T1、T4在内的多种噬菌体的侵染。Southern blot结果显示,该系统抑制了噬菌体DNA在宿主胞内的复制。最后,该研究解析了两个新蛋白SspB与SspE的生化活性:(1)修饰蛋白SspB的结构与核酸酶FokI类似,具有DNA核酸酶活性,可以在DNA上引入单链缺刻;(2)限制蛋白SspE同时具有核酸酶与NTP水解酶活性。有趣的是,SspE的NTP水解酶结构域可以感知携带磷硫酰化修饰的DNA,显示了磷硫酰化限制系统和修饰系统之间的功能耦合。研究者通过分析发现234株细菌基因组携带有该新型磷硫酰化限制修饰系统。
细菌防御系统与噬菌体的对抗与相互进化是生物演化史上精彩的范例。除了限制-修饰系统外,一些细菌的防御系统,如CRISPR-Cas9系统,toxin-antitoxin系统等都在不同领域有所应用。然而近年的研究显示,人们对于这些微生物防御系统的了解还不够全面。该研究鉴定出了以CPSCA单链磷硫酰化修饰为特征的一套全新的细菌防御系统,并揭示了其限制性蛋白SspE是如何感应修饰的功能耦合机理。该研究拓展了磷硫酰化限制-修饰系统的多样性,深化了对细菌防御系统的认识,开拓了磷硫酰化修饰以及细菌防御系统研究新的方向。
另外,DNA除了由ATCG四种碱基组成,构成编码生命信号的遗传物质之外,还有各种各样的表观遗传修饰,在不改变DNA密码的情况下对生命体进行调节。在细菌中,最常见的DNA修饰是由甲基转移酶Dam介导的Gm6ATC甲基化修饰,它可以调控染色体复制的起始,DNA错配修复以及相关基因的表达。
上述工作,武汉大学博士生熊啸林、上海交通大学吴更教授和武汉大学博士生魏月为并列第一作者,王连荣教授为通讯作者。
2020年6月,武汉大学药学院王连荣团队还与合作者在PNAS上发表名为Epigenetic competition reveals density-dependent regulation and target site plasticity of phosphorothioate epigenetics in bacteria的文章,揭示了DNA磷硫酰化修饰的位点选择性及其与DNA三维结构和DNA甲基化修饰的关系。
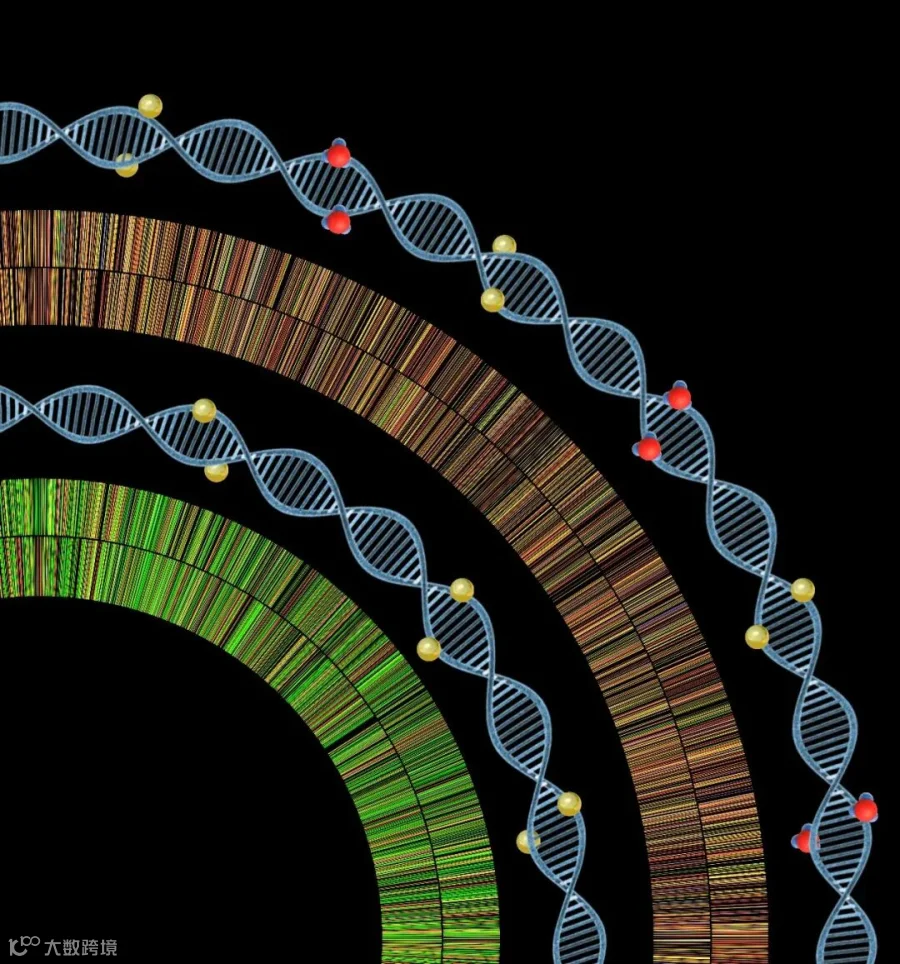
研究者通过染色质免疫沉淀-测序(ChIP-seq)发现沙门氏菌中DNA磷硫酰化修饰蛋白DndACDE主要结合在基因组的GATC序列上,而不是结合在原本的修饰位点GAAC/GTTC上,而GATC序列正是DNA的m6A甲基化序列。通过质谱以及测序技术对含有或敲除甲基化酶Dam的细菌中DNA磷硫酰化修饰位点进行定量及定位分析,研究者发现当敲除甲基化酶Dam后,沙门氏菌中有一半的DNA磷硫酰化修饰位点从GAAC/GTTC序列转移到了GATC序列上。这说明沙门氏菌的磷硫酰化修饰蛋白不仅具有修饰GAAC/GTTC的能力,还同时具有修饰GATC序列的能力。在有甲基化修饰的情况下,GATC序列上的甲基化阻止了Dnd蛋白对该序列进一步的磷硫酰化修饰,从而使沙门氏菌野生型表现出GAAC/GTTC类型的磷硫酰化修饰;而当甲基化缺失之后,Dnd蛋白由于没有甲基化的影响而可以顺利对GATC位点进行磷硫酰化修饰。另外,虽然甲基化缺失导致了磷硫酰化修饰位点发生位移,整个细菌基因组中磷硫酰化修饰的总量保持不变。
图2. 细菌基因组上磷硫酰化修饰位点分布示意图。在有甲基化(红色球)存在,基因组上主要磷硫酰化修饰(金色球)位点为GAAC/GTTC(外圈:桔色和黄色),敲除Dam甲基转移酶基因后,基因组上主要磷硫酰化修饰位点变为GATC(内圈:绿色)。
之后,研究者利用高通量DNA结构预测工具DNAShapeR对已发现的DNA磷硫酰化修饰位点回文序列GNNC进行DNA结构相关的14个参数进行分析,发现整个中心序列可以分为GGCC/GCGC和GAAC/GTTC/GATC/GTAC两个大类,说明不同细菌的Dnd蛋白偏好修饰不同结构的DNA。通过对碱基堆积 (base stacking) 的特性minor groove width (MGW) 和 roll进行分析,发现不同细菌中DNA磷硫酰化修饰序列GAAC/GTTC/GATC/GGCC与其他无关序列GTAC/GCGC表现出明显不同,说明与碱基堆积相关的DNA结构特性更多的决定了DNA的磷硫酰化修饰频率。而通过对含有磷硫酰化修饰细菌的进化树分析,研究者也发现在同一株菌中,其磷硫酰化修饰与甲基化修饰识别的位点往往是不同的。
该研究结合多种生物学以及计算机模拟等技术手段,揭示了DNA磷硫酰化修饰作为一个新型表观遗传修饰的独特性质。DNA结构相似的GAAC/GTTC/GATC序列可以被DNA磷硫酰化修饰蛋白所识别,而GATC上面的甲基化修饰则可以阻碍DNA磷硫酰化系统修饰该位点,使得磷硫酰化修饰位点向GAAC/GTTC位点发生迁移。通常一个细菌中往往含有不止一种DNA修饰系统来行使限制修饰或者表观遗传的功能,本研究阐明了在同一种细菌中两种完全不同类型的DNA修饰之间的相互影响,证明了DNA磷硫酰化修饰的位点特异性与DNA结构的关系,也加深了我们对不同表观修饰系统的相互作用的理解。
上述工作,武汉大学博士研究生武晓琳与曲阜师范大学教授曹博为该论文的并列第一作者,武汉大学为第一作者单位,武汉大学王连荣教授,波士顿大学James E. Galagan教授以及麻省理工学院Peter C. Dedon为共同通讯作者。
https://www.nature.com/articles/s41564-020-0700-6
https://www.pnas.org/content/117/25/14322