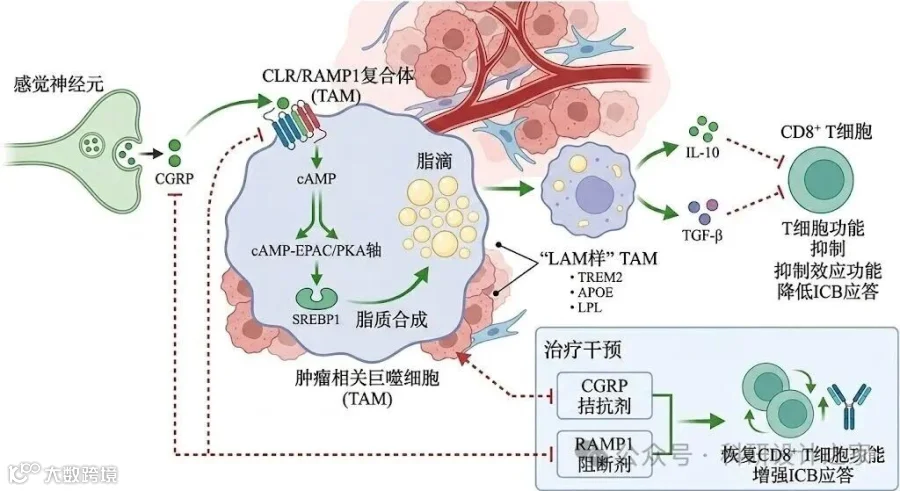
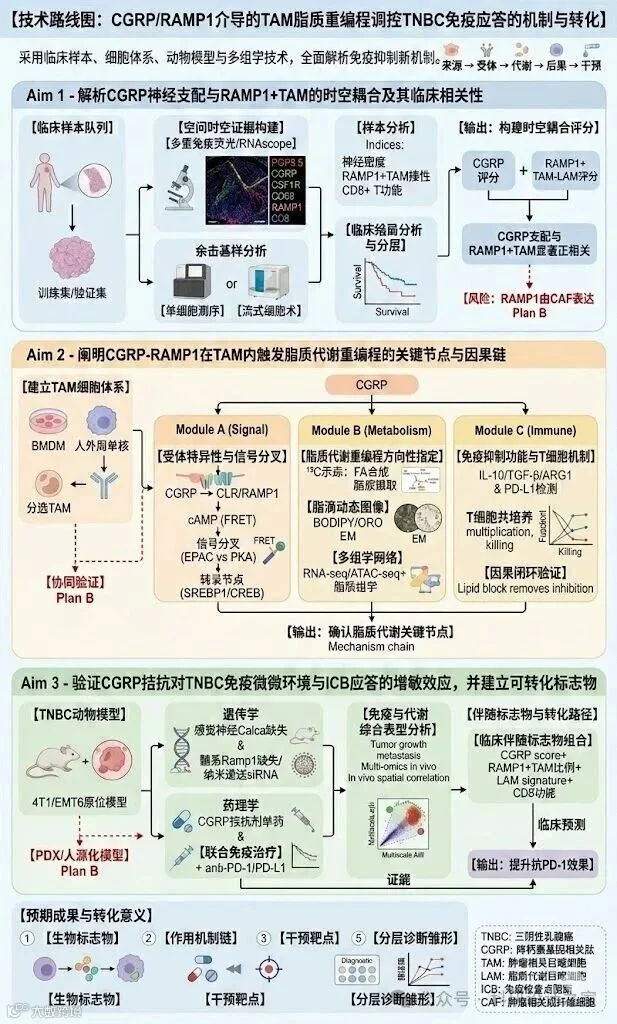
感觉神经肽CGRP通过RAMP1信号驱动肿瘤相关巨噬细胞脂质代谢重编程促进三阴性乳腺癌免疫抑制的机制研究摘要三阴性乳腺癌(TNBC)缺乏明确可药靶,复发转移率高。免疫检查点抑制剂(ICB)虽已进入TNBC治疗体系,但总体获益仍受“免疫排斥/免疫抑制型微环境”制约。近年来“肿瘤神经科学”提示:感觉神经及其神经肽可作为肿瘤微环境的上游调控因子,重塑基质、免疫与代谢网络。我们的前期研究与最新Cell工作表明:TNBC微环境中NGF可激活感觉神经元释放CGRP,并通过RAMP1受体触发下游cAMP信号影响肿瘤微环境结构与免疫浸润。然而,CGRP是否直接作用于肿瘤相关巨噬细胞(TAM),并通过脂质代谢重编程驱动其向“脂质相关巨噬细胞(LAM样)”免疫抑制表型转化,仍缺乏因果证据。本项目提出核心假说:TNBC微环境中感觉神经元来源CGRP与TAM表面CLR/RAMP1复合体结合,激活cAMP-EPAC/PKA轴并启动SREBP1介导的脂质合成/脂滴积累程序,诱导TAM获得TREM2/APOE/LPL等LAM样代谢表型并分泌IL-10、TGF-β等免疫抑制因子,最终抑制CD8+T细胞效应功能与ICB应答。项目将围绕“来源-受体-代谢程序-免疫后果-转化干预”开展三部分研究:①解析TNBC中CGRP神经支配与RAMP1+TAM的时空耦合及其与临床预后的关联;②阐明CGRP-RAMP1在TAM内触发的脂质代谢重编程关键节点与可药靶环节;③评估CGRP拮抗剂/受体阻断与ICB联合在TNBC模型中的增敏效果并建立可转化的生物标志物组合。预期建立“神经肽-免疫代谢-免疫抑制”的新型因果链,为TNBC联合免疫治疗提供可直接推进临床的干预策略与分层指标。 立项依据(一)研究背景:TNBC免疫治疗瓶颈指向“免疫抑制型髓系生态位”三阴性乳腺癌(TNBC)异质性强、可药靶少、复发转移风险高。免疫检查点抑制剂(ICB)联合化疗已在部分TNBC人群中显示获益,但总体有效率仍受限,临床上常见“肿瘤内T细胞稀少/功能受抑”的免疫排斥或免疫抑制型微环境。大量证据提示,肿瘤相关巨噬细胞(TAM)是TNBC免疫抑制网络的枢纽细胞群:其既能通过细胞因子与免疫检查点分子抑制CD8+T细胞,也能通过代谢竞争和代谢产物重塑局部免疫生态。特别是近年“脂质富集/脂滴增多”的TAM亚群被认为与免疫治疗抵抗密切相关,其形成机制与上游驱动信号尚不清晰,是当前TNBC免疫增敏的关键科学空白之一。[17][18](二)前沿进展:肿瘤神经科学提示“神经肽”可作为免疫微环境上游开关肿瘤微环境并非仅由肿瘤细胞、免疫细胞和基质细胞构成;神经系统(感觉/交感/副交感)与肿瘤之间的结构性连接与信号互作,已被提出为肿瘤进展的新维度。综述性研究指出,“神经信号”可与肿瘤代谢、免疫逃逸、血管生成和基质重塑等核心表型交叉,成为影响治疗反应的“可干预变量”。[4][5]在机制层面,最新Cell研究在TNBC场景明确提出:TME中NGF可触发感觉神经元释放CGRP,CGRP与受体RAMP1结合激活cAMP信号并促进致密基质形成,从而驱动免疫排斥;更重要的是,该研究提示CGRP受体拮抗剂可与抗PD-1产生协同,提示“神经肽轴”具备直接的药物转化入口。[1]与此同时,Nature研究显示另一感觉神经肽Substance P可通过“细胞外RNA–TLR7轴”促进转移,为“神经肽-免疫受体”链条提供了可验证范式。[2]此外,NGF–TrkA轴本身也被证明可塑造免疫抑制并影响免疫治疗应答。[3]综合来看,“感觉神经—神经肽—免疫细胞”的因果链正成为CNS级别研究的竞争热点,但在TNBC中是否存在直接作用于髓系细胞代谢程序的关键节点,尚未被系统回答。(三)关键缺口:CGRP是否直接驱动TAM脂质代谢重编程并塑造免疫抑制?CGRP是经典的感觉神经肽,既可通过cAMP依赖的PKA/CREB轴调控免疫细胞转录程序,也可在不同组织环境中呈现“抗炎/促炎”双向效应。[7]免疫学研究提示CGRP可诱导巨噬细胞向调节型表型偏移,促进IL-10产生并抑制过度炎症反应,为其“免疫抑制潜能”提供了直接证据。[8]更具启发性的是,Science Translational Medicine近期在非肿瘤病理环境中证明:伤害感受神经可通过CGRP与巨噬细胞通讯并诱导其呈现免疫抑制样表型,提示“CGRP→巨噬细胞”并非概念推断而是可被实证的细胞间通信轴。[9]另一方面,人源肿瘤样本的单细胞研究显示:肿瘤细胞来源的CGRP也能通过cAMP相关通路影响髓系细胞(如树突状细胞)分化与功能,并可被CGRP受体拮抗剂逆转,提示在肿瘤场景中CGRP具备“非神经元受体靶细胞”的广谱性。[6]因此,提出并验证“感觉神经元来源CGRP直接作用于TAM并触发其代谢重编程”,具有明确的科学逻辑与可验证路径。(四)机制支点:TAM脂质代谢与免疫抑制的耦合,需要上游“触发器”与关键转录节点免疫抑制型TAM常伴随脂滴堆积、脂肪酸合成/摄取增强及脂质信号分子富集,这些代谢特征不仅是表型标志,更可能是维持免疫抑制功能的“代谢必需条件”。已有研究表明,脂滴依赖的脂肪酸代谢可直接控制TAM的免疫抑制表型;当脂滴代谢被干预,TAM可发生功能性重塑并解除对T细胞的抑制。[12]在肿瘤免疫生态位中,Treg等免疫细胞可通过改变局部细胞因子谱与代谢约束,促进肿瘤促生长型巨噬细胞的脂质代谢适配,其中SREBP1等脂质代谢转录程序被认为是关键枢纽。[10]此外,TREM2等“脂质相关髓系标志物”在多癌种中被证实与免疫抑制型TAM富集相关,针对TREM2的抗体干预可重塑TAM状态并增强抗肿瘤免疫反应,提示“LAM样TAM”具备明确的可药性方向。[13]而CD36等脂质摄取通路亦被证实可驱动转移相关巨噬细胞的脂质富集与促转移功能,为“脂质摄取/合成—免疫抑制”提供补充证据。[14]综上,TAM脂质代谢与免疫抑制的耦合已经被多条证据链支持,但“是什么上游信号在TNBC中启动这一代谢程序”仍是空缺。我们提出CGRP-RAMP1正是这一缺口的候选触发器。(五)研究基础与可行性:多组学数据支持“RAMP1+髓系亚群—脂质代谢—不良预后”的关联框架公共单细胞转录组数据为本项目提供了可复现的证据来源。TNBC单细胞研究(含GEO数据集)揭示了肿瘤髓系细胞存在明显的功能与代谢异质性,为筛查“受体阳性且脂质代谢富集”的TAM亚群提供了数据基础。[15]TCGA-BRCA的分子分型与生存数据可用于验证RAMP1/相关通路高表达与TNBC不良预后的关联。[17]CPTAC等蛋白质组学资源可进一步支持“受体轴—脂质合成酶(如FASN)—免疫抑制标志”在蛋白水平的协同变化,为后续机制验证与转化标志物筛选提供更高维度证据。[16]与此同时,前沿综述指出肿瘤神经系统与免疫系统互作具有显著“双向性”:神经肽影响髓系细胞命运的同时,免疫细胞也可通过分泌神经营养相关因子、轴突导向分子及炎症介质反向塑造肿瘤神经支配与疼痛回路。这为我们进一步探索“CGRP驱动TAM→TAM反向维持神经-免疫正反馈”的扩展假说提供了合理生物学基础。[11](六)拟解决的关键科学问题与总体目标围绕“神经肽→受体→代谢程序→免疫后果”主线,本项目拟回答三类可检验问题:1)TNBC中CGRP神经支配与RAMP1阳性TAM是否存在稳定的时空耦合?其与T细胞功能抑制、免疫排斥及预后不良之间是否存在统计学与空间组织学的一致性?2)CGRP-RAMP1在TAM内触发的关键代谢程序是什么:以脂肪酸合成为主还是以脂质摄取/脂滴代谢为主?其关键转录节点(如SREBP1等)与信号分叉(EPAC/PKA)能否被明确定位并实现因果证据闭环?3)利用已具备临床可及性的CGRP受体拮抗策略,是否能在TNBC中“可重复地”重塑TAM代谢与免疫抑制,并显著提高ICB应答?能否建立可转化的联合用药人群分层标志物?项目总体目标:阐明CGRP-RAMP1轴驱动TAM脂质代谢重编程促进TNBC免疫抑制的分子机制,并提出可转化的“CGRP拮抗剂+ICB”联合增敏策略,为TNBC免疫治疗提供新的可药靶与分层指标。(七)创新点凝练(面上项目“可验证创新”口径)创新点1(新机制链):提出并验证“感觉神经肽CGRP作为TAM脂质代谢重编程的上游触发器”,将肿瘤神经科学与免疫代谢学在TNBC语境下做实证耦合,区别于既往以CAF/基质为主的CGRP研究路径。[1][4][5]创新点2(新靶点组合):以RAMP1为入口,解析EPAC/PKA分叉与脂质代谢转录程序的关键节点,形成“可药靶-可读出-可分层”的干预闭环,并将TREM2/CD36等LAM样表型标志纳入机制链条验证与风险替代路径。[10][13][14]创新点3(新转化策略):借助临床可及的CGRP受体拮抗剂(小分子拮抗剂与单抗均已在其他适应证获批),在TNBC模型中系统评估其与ICB的协同增敏并构建伴随诊断指标,显著降低从机制到临床的转化门槛。[1][6][19][20]研究内容一、总体思路与技术路线(文字版)围绕“来源—受体—代谢程序—免疫后果—转化干预”五段式闭环,本项目采用“人群证据(临床样本与多组学)+ 因果验证(遗传学/药理学)+ 机制拆解(代谢通量与转录调控)+ 转化评估(联合免疫治疗)”的组合策略。总体上先建立关联与空间证据,再进行受体特异性的因果敲定,最后以联合治疗与伴随标志物输出实现面上项目应有的闭环交付。二、研究内容1:解析TNBC中CGRP神经支配与RAMP1+TAM的时空耦合及其临床相关性1.1 目标与关键指标目标:在“组织空间—单细胞分群—临床结局”三层面确认CGRP与RAMP1+TAM的耦合关系,并锁定与免疫抑制/免疫排斥相关的可量化读出。关键指标:①CGRP+神经纤维密度(PGP9.5/CGRP共染);②RAMP1+TAM比例与空间邻近度(RAMP1/CSF1R或CD68,并联合TREM2/APOE/LPL等LAM标志);③CD8+T细胞密度与功能评分(GZMB/IFNG或多重免疫荧光);④临床结局(复发/无进展生存)与分层相关性。1.2 研究设计与方法学要点(1)临床样本队列:收集TNBC手术样本与配对临床信息,建立“训练集+验证集”的分层分析框架;必要时引入公开空间转录组数据作为外部验证。(2)空间证据构建:采用多重免疫荧光/免疫组化与RNAscope结合,定量“神经纤维—TAM—T细胞”三者空间关系;使用空间邻近分析(nearest-neighbor / Ripley’s K等)评价耦合强度。(3)单细胞与蛋白水平验证:在部分样本开展scRNA-seq/CITE-seq或流式细胞术,确认RAMP1在TAM亚群中的表达与LAM样代谢签名共现;并以蛋白水平(流式/质谱/免疫印迹)确认受体与脂质代谢酶表达。(4)输出:构建“CGRP神经支配评分 + RAMP1+TAM-LAM评分”的联合分层指标,用于后续Aim3的伴随诊断雏形。1.3 预期结果与判定预期在TNBC中观察到:CGRP神经支配增强区域富集RAMP1+TAM,并与CD8+T细胞稀少/功能低下呈负相关;RAMP1+TAM具有更高的脂滴/脂质代谢签名。若仅观察到RAMP1主要表达于CAF而非TAM,则将该结果作为“风险分支触发信号”,转入Aim2/替代策略中验证“间接通路(CGRP→CAF→TAM)”的贡献。三、研究内容2:阐明CGRP-RAMP1在TAM内触发脂质代谢重编程的关键节点与因果链2.1 目标与核心问题目标:回答“CGRP是否直接驱动TAM脂质代谢重编程”以及“关键转录/代谢节点是什么”。在可验证范围内把信号链条拆解为:受体特异性(CLR/RAMP1)→第二信使(cAMP)→信号分叉(EPAC与/或PKA)→转录节点(以SREBP1为核心候选,同时评估CREB等)→脂质合成/摄取/脂滴动态→免疫抑制输出。2.2 关键实验模块模块A:受体特异性与信号分叉定位(1)细胞体系:使用骨髓来源巨噬细胞(BMDM)、单核细胞来源巨噬细胞(人外周血)、以及从TNBC肿瘤组织分选获得的原位TAM;并设置肿瘤条件培养基/乳酸/低氧等TME模拟条件。(2)受体干预:采用RAMP1/CALCRL敲降或条件性敲除(或CRISPR编辑)验证CGRP效应的受体依赖性;并用受体拮抗剂进行药理学互证。(3)信号读出:实时检测cAMP变化(FRET探针/ELISA),并区分EPAC与PKA分叉(Rap1-GTP拉下、PKA底物磷酸化、CREB磷酸化);使用特异性抑制剂/激动剂进行因果定位。模块B:脂质代谢重编程的方向性鉴定(合成 vs 摄取 vs 脂滴代谢)(1)脂滴动态:BODIPY/油红O染色,结合定量成像分析脂滴数量、体积与分布;并通过电镜或共聚焦验证脂滴-线粒体接触变化。(2)代谢通量:开展13C-葡萄糖/13C-乙酸示踪,解析脂肪酸新生合成贡献;同时评估脂质摄取(荧光标记脂肪酸/LDL摄取)与关键转运体表达。(3)多组学:RNA-seq/ATAC-seq结合脂质组学,构建“信号—转录—代谢产物”网络图谱,锁定最稳定、最可药的节点组合。模块C:免疫抑制功能输出与T细胞抑制机制(1)细胞因子与检查点:检测IL-10、TGF-β、ARG1等免疫抑制因子及PD-L1等分子;建立“代谢读出—免疫抑制读出”的定量关联。(2)功能评估:TAM与CD8+T细胞共培养,检测T细胞增殖、细胞毒分子表达与杀伤能力;区分可溶性因子抑制与细胞接触抑制。(3)因果闭环:在阻断脂质代谢关键步骤后,观察免疫抑制是否被解除,从而证明“脂质代谢重编程是免疫抑制的必要条件”。2.3 预期结果与判定预期CGRP可在受体依赖条件下提升TAM脂滴积累并诱导LAM样转录签名,同时增强其对CD8+T细胞的抑制作用;抑制关键分叉或代谢节点将显著逆转上述效应。若CGRP在纯巨噬细胞体系中效应弱,则将重点转入“细胞互作增强”情境:加入CAF/肿瘤细胞建立三元共培养与条件培养基互换,以判定是否存在“间接驱动+协同放大”的真实生物学模式。四、研究内容3:验证CGRP拮抗策略对TNBC免疫微环境与ICB应答的增敏效应,并建立可转化分层标志物3.1 目标在体内模型中证明:阻断CGRP-RAMP1轴可重塑TAM脂质代谢与免疫抑制,从而提升抗PD-1/PD-L1治疗反应;同时输出“可分层、可检测”的伴随标志物组合。3.2 研究设计(1)动物模型与遗传学验证a)建立同系原位TNBC模型(如4T1/EMT6),评估肿瘤生长、肺转移与生存。b)感觉神经源性CGRP验证:采用感觉神经元特异性Calca缺失或感觉神经抑制策略,证明上游来源的必要性。c)髓系受体依赖验证:构建髓系细胞特异性Ramp1缺失或采用TAM定向递送的基因干预策略(如靶向巨噬细胞的纳米递送siRNA/CRISPR元件),验证“受体在TAM上”的必要性。(2)药理学干预与联合免疫治疗a)单药:CGRP受体拮抗策略(小分子/抗体/肽段)分别评估对TAM脂滴、LAM样签名与免疫浸润的影响。b)联合:在明确安全剂量窗内,与抗PD-1/PD-L1联用,比较肿瘤控制率、免疫浸润质量与记忆T细胞形成;重点回答“是否存在明确协同”以及“协同依赖的机制节点”。(3)免疫与代谢综合表型分析a)免疫谱:流式/质谱流式/单细胞测序评估TAM、T细胞、树突状细胞等组成与状态变化。b)代谢谱:肿瘤组织脂质组学与免疫细胞分选后的代谢组学,验证“体外机制节点”在体内的可复现性。c)空间验证:在体内样本中复现Aim1的空间耦合分析,形成“机制—空间—疗效”三维证据链。3.3 伴随标志物与转化路径(1)标志物组合:CGRP神经支配评分 + RAMP1+TAM比例 + LAM样代谢签名(TREM2/APOE/LPL等) + CD8功能评分,形成风险分层与疗效预测模型。(2)可检测性:优先选择免疫组化/多重免疫荧光可实现的标志物作为临床可落地版本;同时探索血浆CGRP水平与外周髓系细胞受体表达作为“低侵入”读出。(3)转化可行性:利用既有药物安全性信息与免疫治疗临床路径,将“CGRP拮抗剂+ICB”定位为可快速推进的联合策略,为后续临床研究奠定基础。 3.4 预期结果与判定预期阻断CGRP-RAMP1轴可降低TAM脂滴与LAM样签名,增强CD8+T细胞浸润与效应功能,并显著提高ICB疗效;伴随标志物可用于分层预测。若联合疗效不显著,则基于Aim2机制结果调整:选择更精准的代谢节点联合(如脂质摄取/脂滴代谢关键步骤)或在更符合临床的模型(PDX/人源化)中验证。 五、年度计划与关键里程碑(3年)第1年:完成TNBC样本队列建立与空间/单细胞验证;形成CGRP神经支配与RAMP1+TAM耦合的量化指标;建立体外巨噬细胞体系与受体依赖性验证。第2年:完成信号分叉与代谢通量拆解,锁定1–2个关键可药节点;完成T细胞抑制机制的必要性验证;形成可复现的“机制闭环”数据包。第3年:完成遗传学与药理学体内验证;完成联合ICB增敏评估与伴随标志物模型;输出可转化的治疗策略与分层指标,并形成高质量论文与后续临床转化方案。六、风险点与替代策略(评审关注项前置回答)风险1:RAMP1在TAM表达较低或主要集中于CAF。替代:采用高灵敏空间与蛋白检测确认真实表达;若确为CAF主导,则转向“CGRP→CAF→代谢因子/细胞外基质→TAM脂质重编程”的间接通路,并以细胞互作体系完成因果闭环;同时评估其他RAMP家族成员或相关受体复合体作为替代受体入口。风险2:CGRP诱导脂滴增加但与免疫抑制不构成必要关系。替代:引入“必要性”实验(阻断脂滴生成/脂质代谢关键步骤)与“充分性”实验(人工激活代谢节点)来严格界定因果;若必要性不成立,则将免疫抑制机制转向cAMP下游的细胞因子调控或抗原呈递缺陷等方向。风险3:药理学拮抗剂在动物模型中的效应不稳定。替代:采用多种拮抗策略交叉验证(小分子、抗体、肽段与遗传学缺失);并优化给药窗口与联合方案,必要时采用肿瘤靶向递送提高局部有效暴露。七、预期成果(1)科学产出:提出并验证TNBC中“CGRP-RAMP1驱动TAM免疫代谢重编程”的新机制链;形成可被独立重复的空间-单细胞-因果证据体系。(2)应用产出:形成“CGRP拮抗剂+ICB”联合增敏策略的前临床证据;建立可落地的伴随诊断分层指标组合。(3)人才与平台:沉淀TNBC神经-免疫-代谢多组学分析流程与空间免疫代谢评估体系,为后续系列项目与临床转化研究提供平台支撑。参考文献(共20篇,仅在立项依据中引用标注)[1] Zhang SW, Wang H, Xiao Y, et al. Sensory neurons drive immune exclusion by stimulating a dense extracellular matrix in the breast cancer tumor microenvironment. Cell. 2026;189(4):1039-1055.e20. doi:10.1016/j.cell.2026.01.001.[2] Padmanaban V, Keller I, Seltzer ES, et al. Neuronal substance P drives metastasis through an extracellular RNA–TLR7 axis. Nature. 2024;633:207–215. doi:10.1038/s41586-024-07767-5.[3] Yin T, Wang G, Wang L, et al. Breaking NGF–TrkA immunosuppression in melanoma sensitizes immunotherapy for durable memory T cell protection. Nature Immunology. 2024;25:268–281. doi:10.1038/s41590-023-01723-7.[4] Mancusi R, Monje M. The neuroscience of cancer. Nature. 2023;618:467–479. doi:10.1038/s41586-023-05968-y.[5] Hanahan D, Monje M. Cancer hallmarks intersect with neuroscience in the tumor microenvironment. Cancer Cell. 2023;41(3):573–580. doi:10.1016/j.ccell.2023.02.012.[6] Hou Y, et al. The neurotransmitter calcitonin gene-related peptide shapes an immunosuppressive microenvironment in medullary thyroid cancer. Nature Communications. 2024;15:5555. doi:10.1038/s41467-024-49824-7.[7] Baral P, Udit S, Chiu IM. Pain and immunity: implications for host defence. Nature Reviews Immunology. 2019;19:433–447. doi:10.1038/s41577-019-0147-2.[8] Baliu-Piqué M, Jusek G, Holzmann B. Neuroimmunological communication via CGRP promotes the development of a regulatory phenotype in macrophages. European Journal of Immunology. 2014;44(12):3708–3716. doi:10.1002/eji.201444546.[9] Fattori V, et al. Nociceptor-to-macrophage communication through CGRP promotes endometriosis by inducing an immunosuppressive macrophage phenotype. Science Translational Medicine. 2024;16(737):eadk7598. doi:10.1126/scitranslmed.adk7598.[10] Liu C, et al. Regulatory T cells promote SREBP1-dependent metabolic fitness of tumor-promoting macrophages via repressing CD8+ T cell-derived interferon-γ. Immunity. 2019;51(3):560–574.e6. doi:10.1016/j.immuni.2019.08.018.[11] Pu Y, et al. Neuro-immune crosstalk in cancer. Cell Discovery. 2025. (综述)[12] Wu H, et al. Lipid droplet-dependent fatty acid metabolism controls the immunosuppressive phenotype of tumor-associated macrophages. EMBO Molecular Medicine. 2019. (原始研究)[13] Binnewies M, et al. Anti-TREM2 (or genetic Trem2 loss) reprograms tumor-associated macrophages and enhances antitumor immunity. Cell Reports. 2021;37(8):109844. doi:10.1016/j.celrep.2021.109844.[14] Yang P, et al. CD36-mediated fatty acid uptake programs metastasis-associated macrophages and promotes breast cancer metastasis. Nature Communications. 2022;13:5916. doi:10.1038/s41467-022-33349-y.[15] Cristea S, et al. Genetic determinants of gene expression in TNBC at single-cell resolution (GEO: GSE118389). Nature Communications. 2018. PMID:30181541.[16] Krug K, et al. Proteogenomic landscape of breast cancer (CPTAC). Cell. 2020.[17] The Cancer Genome Atlas Network. Comprehensive molecular portraits of human breast tumours. Nature. 2012;490:61–70. doi:10.1038/nature11412.[18] Schmid P, et al. Atezolizumab and nab-paclitaxel in advanced triple-negative breast cancer (IMpassion130). New England Journal of Medicine. 2018. doi:10.1056/NEJMoa1809615.[19] Pfizer. Nurtec ODT (rimegepant) prescribing information (FDA label). Revised 2025; initial FDA approval 2020.[20] U.S. FDA / Amgen. Aimovig (erenumab-aooe) prescribing information (FDA label). First FDA-approved CGRP pathway monoclonal antibody for migraine prevention, 2018.





 生物医学AI圈子
生物医学AI圈子