
2026-02-21
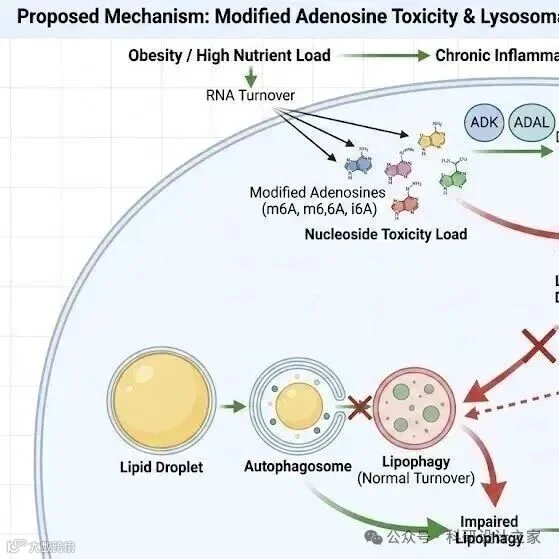
2026-02-21

2026-02-19
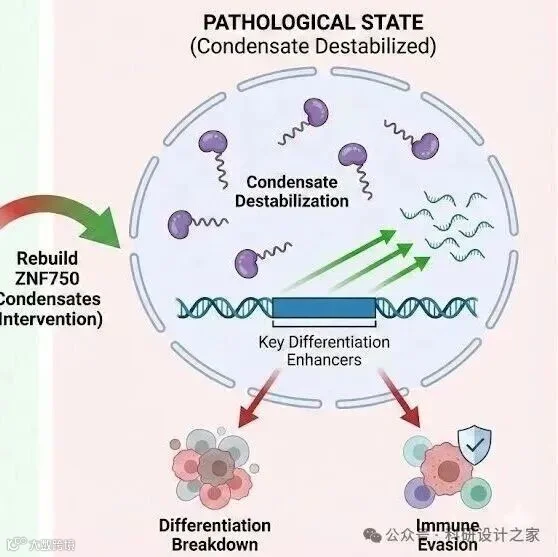
2026-02-19
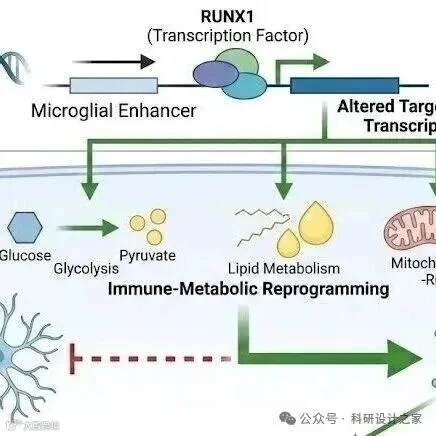
2026-02-19

2026-02-18
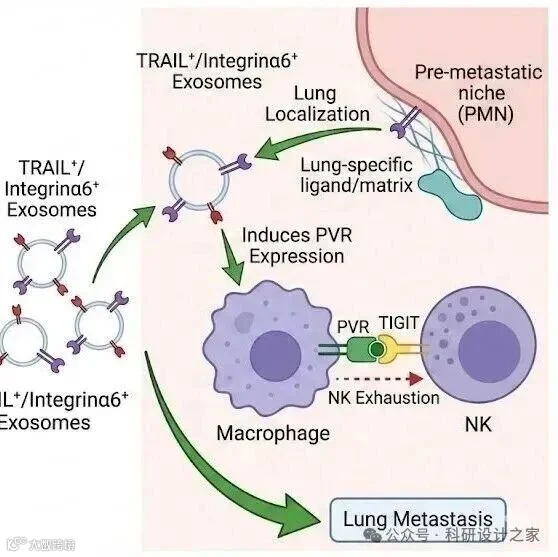
2026-02-18

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

 生物医学AI圈子
生物医学AI圈子
2026-02-21
2026-02-21
2026-02-19
2026-02-19
2026-02-19
2026-02-18
2026-02-18
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|