2026-03-12
2026-03-12

2026-03-11
2026-03-11
2026-03-08
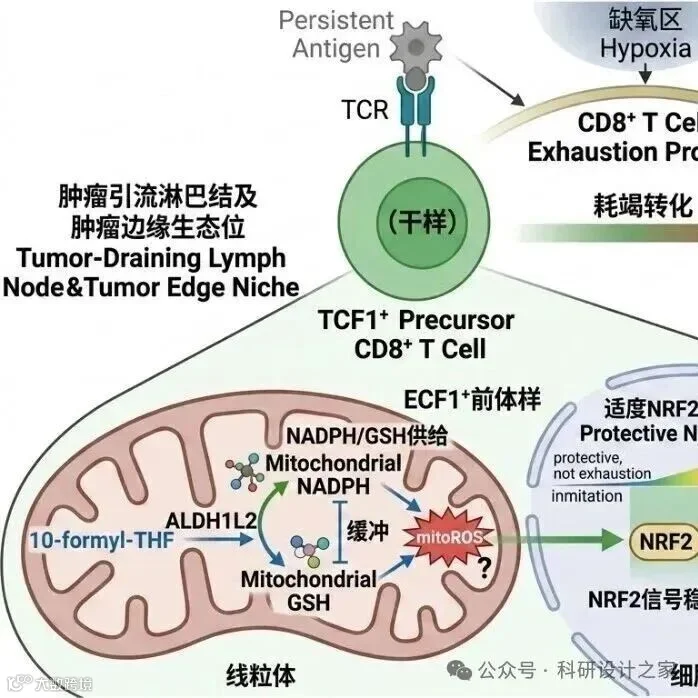
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
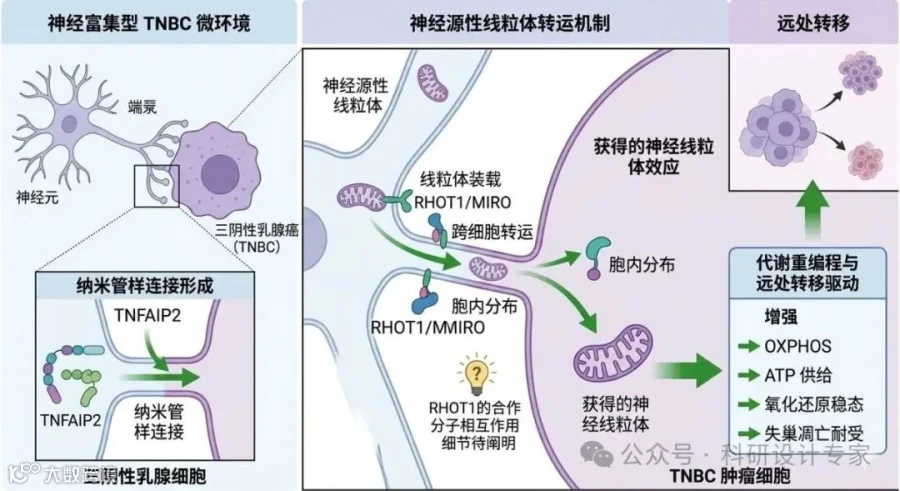
(二)神经-肿瘤共培养及线粒体示踪。建立DRG神经元或iPSC来源感觉神经元与MDA-MB-231、BT-549、4T1等TNBC细胞的直接共培养体系。通过mito-Dendra2、mito-GFP、MitoTracker、mtDNA特异标记及永久示踪策略追踪神经线粒体的跨细胞转运,并采用共聚焦、流式和高内涵系统进行定量。
(三)分子机制研究。采用CRISPR/Cas9、shRNA、过表达和结构域救援策略分别调控TNFAIP2和RHOT1;检测纳米管样连接形成、F-actin重构、RAC1活化、IQGAP1/ITGB4复合体形成及外源线粒体进入后的亚细胞分布;必要时进一步评估TRAK/KIF/MYO相关运动装置在RHOT1依赖运输中的作用。
(四)功能研究。利用Seahorse检测OCR/ECAR,结合ATP、ROS、膜电位、mtDNA拷贝数、球形成、失巢凋亡、迁移侵袭、剪切力耐受及内皮穿越实验,评估神经线粒体输入后的代谢和恶性表型变化。
(五)体内验证。构建乳腺脂肪垫原位移植模型和尾静脉/心内注射模型,结合神经示踪或神经调控手段观察转移灶形成;对RHOT1/TNFAIP2进行遗传或药理干预,评价对原发瘤生长、肺/脑转移和生存的影响。
风险二:TNT样结构并非唯一传递方式。对策:同步评估细胞外囊泡、细胞融合或吞饮等替代途径;若发现多种途径并存,将重点分析TNFAIP2/RHOT1在其中的共性和特异作用。
风险三:RHOT1或TNFAIP2单独干预效应有限。对策:采用双基因联合干预、关键结构域救援和上下游通路联合阻断,界定真正的限制环节。
风险四:体内模型中神经变量复杂。对策:在保留原位微环境的同时,增加实验性转移模型、离体共培养和组织切片验证,以提高证据稳健性。
风险五:转运事件与转移之间因果性不足。对策:把“是否获得神经线粒体”作为谱系分层变量,比较获得组与未获得组的转移能力和代谢特征,从而增强因果推断。
第2年:完成TNFAIP2与RHOT1在连接形成和线粒体转运中的机制研究,形成较完整的分子作用网络。
第3年:完成代谢与转移功能验证、体内干预评价和关键结论整合,形成可发表的高质量论文和可申报后续基金/专利的候选靶点。
预期成果包括:发表相关论文2-3篇;形成神经富集型TNBC与RHOT1/TNFAIP2高表达相关的证据;建立神经源性线粒体转运评价体系;提出可用于转移风险分层或干预的新机制模型。