2026-03-12
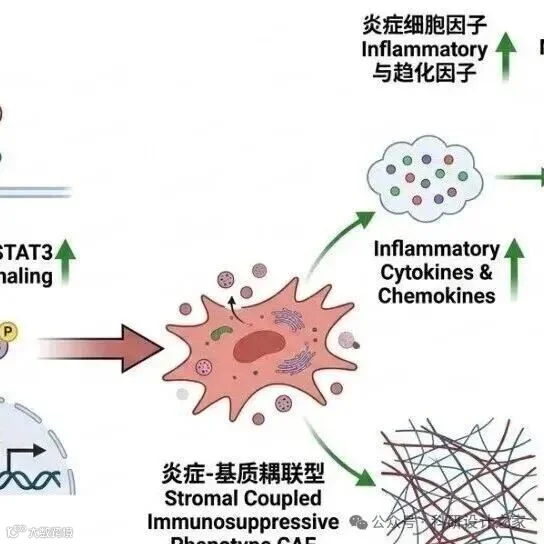
2026-03-12

2026-03-11
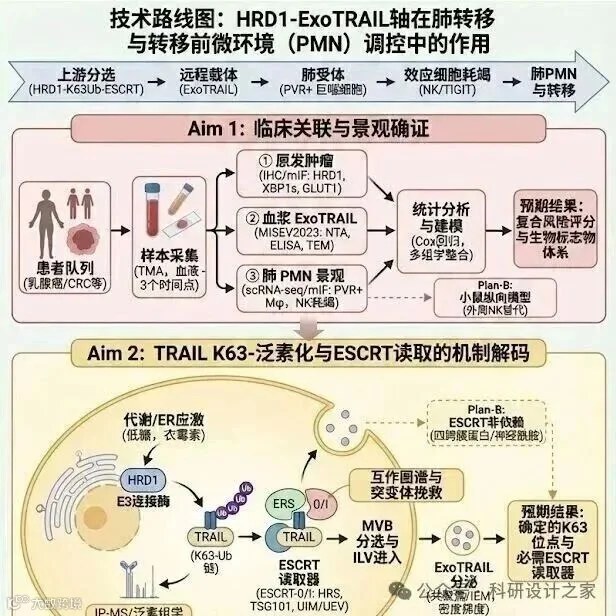
2026-03-11
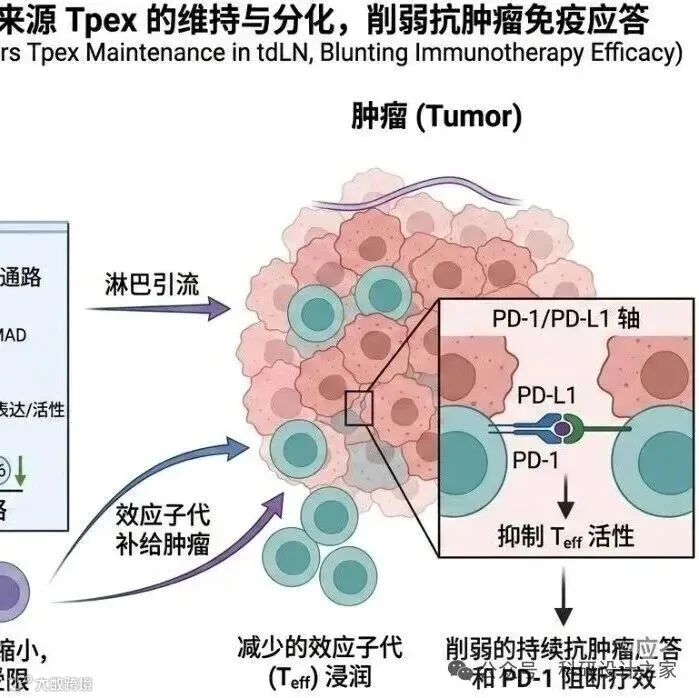
2026-03-08
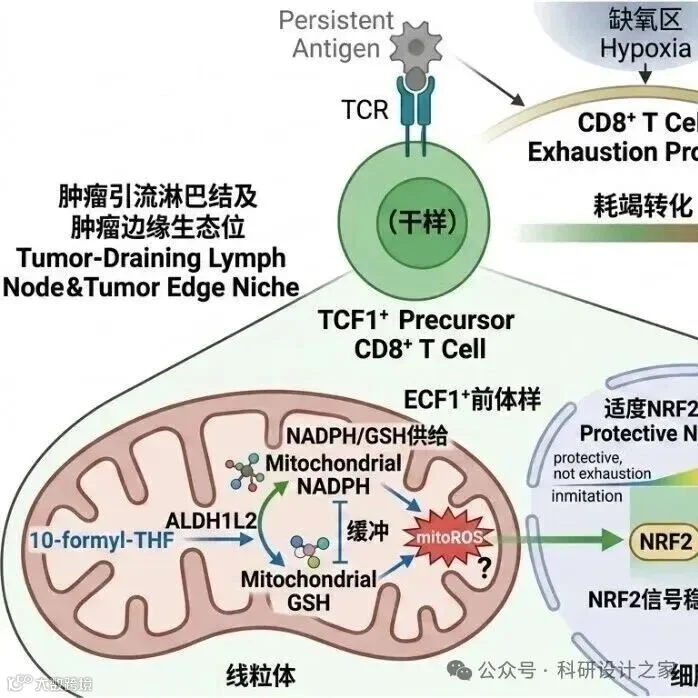
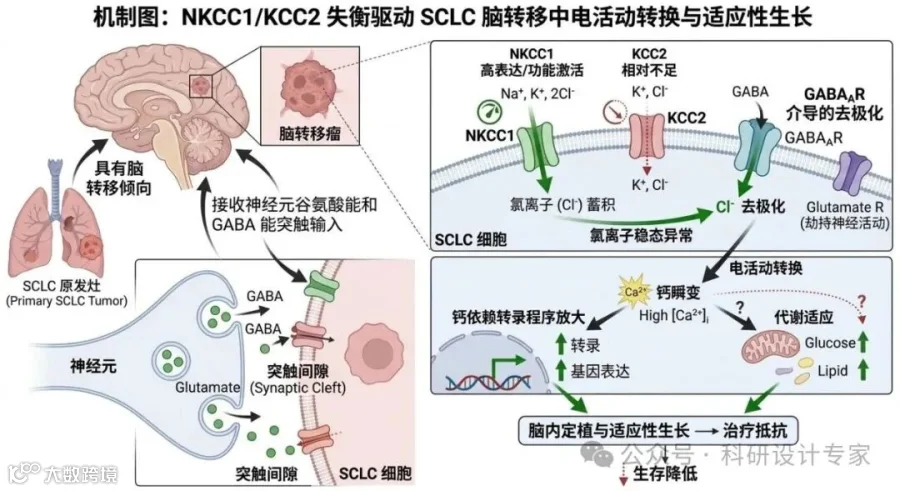
驱动小细胞肺癌脑内适应性生长的机制研究
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
目标二:阐明NKCC1如何决定GABAA信号方向,并驱动SCLC的膜去极化、钙瞬变、转录和代谢重编程。
目标三:验证阻断NKCC1对SCLC脑内定植、生长与治疗反应的影响,评价其联合干预潜力。
2)在本中心原发灶与脑转移灶样本中,采用IHC、多重免疫荧光、RNAscope和空间转录组,解析NKCC1/KCC2与突触标志物、Ca2+信号分子、增殖标志物及胶质细胞相互作用的空间耦联。
3)基于临床队列评估NKCC1高表达与脑转移发生、进展速度、治疗反应和生存的关系,构建预后分层模型。
拟解决的问题是:NKCC1/KCC2失衡是否是SCLC脑适应与脑转移的稳定分子基础。
2)通过遗传学手段(CRISPR敲低/敲除、过表达、救援)和药理学手段(NKCC1抑制剂)操纵NKCC1/KCC2平衡,比较其对GABAA输入方向、动作电位样放电、ERK/CREB/NFAT等下游通路以及细胞增殖、迁移和克隆形成的影响。
3)在与感觉神经元、皮层神经元或星形胶质细胞共培养体系中,观察神经输入对NKCC1依赖性表型的放大效应,并结合转录组和代谢组分析明确其对氧化磷酸化、乳酸利用和脑内存活程序的调节。
拟解决的问题是:SCLC如何把神经网络中的抑制性语言“翻译”为促生长生物电语言。
2)比较单独阻断NKCC1、阻断谷氨酸输入、双重阻断以及与放疗/标准治疗联用的疗效差异,评估其对脑内适应性生长的抑制强度。
3)分析治疗后肿瘤细胞转录状态、突触相关基因表达、胶质细胞反应和免疫微环境变化,明确疗效改善是来源于“电生理脱耦”“代谢失配”还是“脑生态位重塑”。
拟解决的问题是:NKCC1是否能够作为SCLC脑转移的真正可药物化弱点。
第二步,完成细胞和类器官层面的因果验证。通过电生理、氯离子成像、钙成像和遗传干预,直接证明NKCC1控制GABA信号方向。
第三步,完成神经共培养和脑转移模型验证。解析来自不同神经元亚群的输入在NKCC1依赖背景下如何影响SCLC生长。
第四步,完成治疗学验证。以NKCC1阻断为核心,探索其与神经输入阻断或标准治疗协同的机制基础和最佳组合。
(2)分子层面:CREB、pERK、NFAT、OXPHOS相关分子和突触基因变化,作为“神经输入被转录程序接收”的判据。
(3)表型层面:脑内定植率、病灶数量、体积、生存时间及神经症状评分,作为“脑内适应性生长被抑制”的判据。
(4)联合治疗层面:与谷氨酸输入阻断、放疗或标准治疗联用后获得明显优于单药的效果,作为“存在转化潜力”的判据。
统计学方面,细胞实验至少三次独立重复;动物实验采用随机化和盲法;多组比较用ANOVA,生存分析用Kaplan-Meier和Cox模型;组学分析采用FDR校正,确保结论稳健。
**创新点二:把脑转移生态位中的电生理适应纳入SCLC研究主轴。** 既往SCLC脑转移研究多关注胶质细胞支持、代谢适应和血脑屏障穿越,本项目将“生物电适应”置于同等重要地位,有望形成新范式。
**创新点三:构建“突触输入—离子稳态—钙信号—代谢/转录—脑内生长”的完整链条。** 该链条兼顾神经科学严谨性与肿瘤生物学可解释性,具有较强理论整合价值。
**创新点四:靶点可药物化、验证路径清晰。** NKCC1并非抽象概念,而是可通过遗传和药理双路径干预的实际节点,便于后续临床转化。
第二,关键模型成熟。SCLC细胞系、患者来源样本、脑转移模型、神经元共培养、膜片钳和氯离子/钙成像均具有可复制的技术基础;配合类器官和空间转录组,可实现从机制到生态位的跨尺度验证。
第三,指标体系可量化。项目将以反转电位、胞内氯浓度、Ca2+峰值、转录状态、脑内定植率和生存获益作为多层级读出,具有高度可操作性。
第四,转化出口明确。若NKCC1阻断显示显著疗效,可进一步延伸到与突触信号阻断、放疗或标准治疗的联合策略;若某些药物脑穿透不足,也可通过局部递送或新型抑制剂继续推进。
**第二年:** 完成NKCC1决定去极化GABA信号的分子与功能验证,明确其对Ca2+依赖转录/代谢程序的影响。
**第三年:** 完成脑转移模型与联合治疗实验,构建完整机制闭环并形成高水平论文。
预期结果包括:确立NKCC1介导的氯离子稳态异常是SCLC脑内适应性生长的重要基础;提出SCLC脑转移的新型神经电生理机制;为基于离子稳态或突触信号阻断的干预策略提供实验依据。
从评审偏好看,SCLC课题最容易被质疑之处在于“脑内模型复杂、机制链过长”。因此,正式文本中宜强调本项目的主线高度收束:只围绕NKCC1如何决定GABA信号方向这一核心问题展开,并将谷氨酸输入、胶质支持、脑转移生态位等内容定位为验证背景或增强模块,而不喧宾夺主。这样既能保持问题聚焦,又能体现项目对领域前沿的整合能力。
同时,建议在申请书‘预期成果’中明确提出:本项目预期形成1套SCLC去极化GABA电生理判读流程、1套脑转移相关NKCC1/KCC2空间评估指标,以及1个可用于后续药物组合筛选的神经元—SCLC共培养平台。把平台性产出写清楚,有利于增强面上项目的持续产出感和评审认可度。整体上,这将使项目从单篇文献延伸为一条可持续拓展的研究路线。并为后续转化奠基。