

p53信号通路
Section.01
p53基因、蛋白结构与生物学作用
1.TP53基因与p53蛋白结构
TP53基因定位于17p13.1,编码393个氨基酸的p53蛋白。p53包含五个高度保守的功能结构域:N端转录激活结构域(TAD,含TAD1和TAD2)、富含脯氨酸结构域(PRD)、中央DNA结合结构域(DBD)、寡聚化结构域(TD)和C端调节结构域(CTD)[1][2]。其中DBD是核心区域,约80%的突变发生于此。 锌离子对稳定DBD结构至关重要,锌缺乏会导致蛋白热稳定性下降及聚集倾向增加[2]
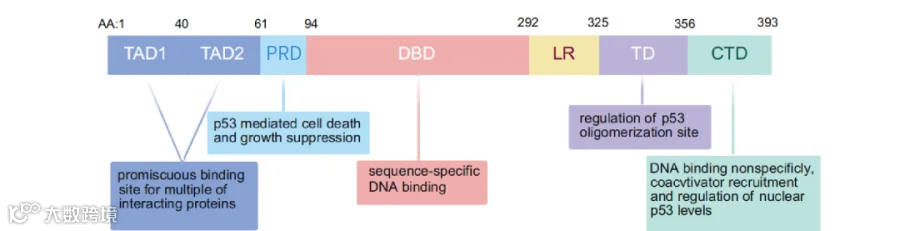
图1. p53蛋白的主要结构域及功能

1.TP53突变类型与功能
TP53突变在人类肿瘤中极为常见,且约70%为错义突变[1]。根据对p53功能的影响,TP53突变可分为三类:
功能缺失型突变(loss of function, LOF):p53丧失抑癌功能
显性负效应突变(dominant-negative effect, DNE):突变蛋白与野生型p53形成混合四聚体,干扰野生型p53功能。
功能获得型突变(gain of function, GOF):突变p53蛋白不仅失去抑癌功能,还获得促进肿瘤进展、转移、治疗抵抗和免疫逃逸的新功能。
TP53六个热点突变残基均位于DNA结合结构域,其中R175H、R248Q、R248W、R273H、R282W等是研究最为广泛的突变类型[1]。不同组织器官中TP53突变谱存在差异——在卵巢癌(47.27%)、结直肠癌(44.55%)、肺癌(40.8%)中突变率较高,而在甲状腺癌(11.13%)、肾癌(8.75%)中突变率较低。
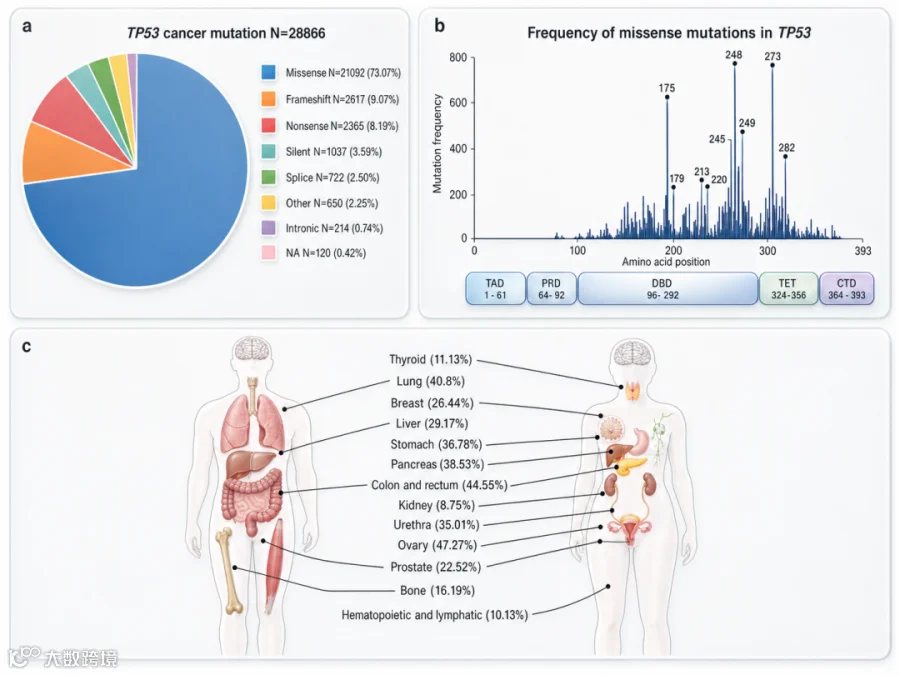
图2. 肿瘤中的TP53基因突变[1]
Section.02
p53信号转导机制:上游调控与下游效应
1.上游调控机制
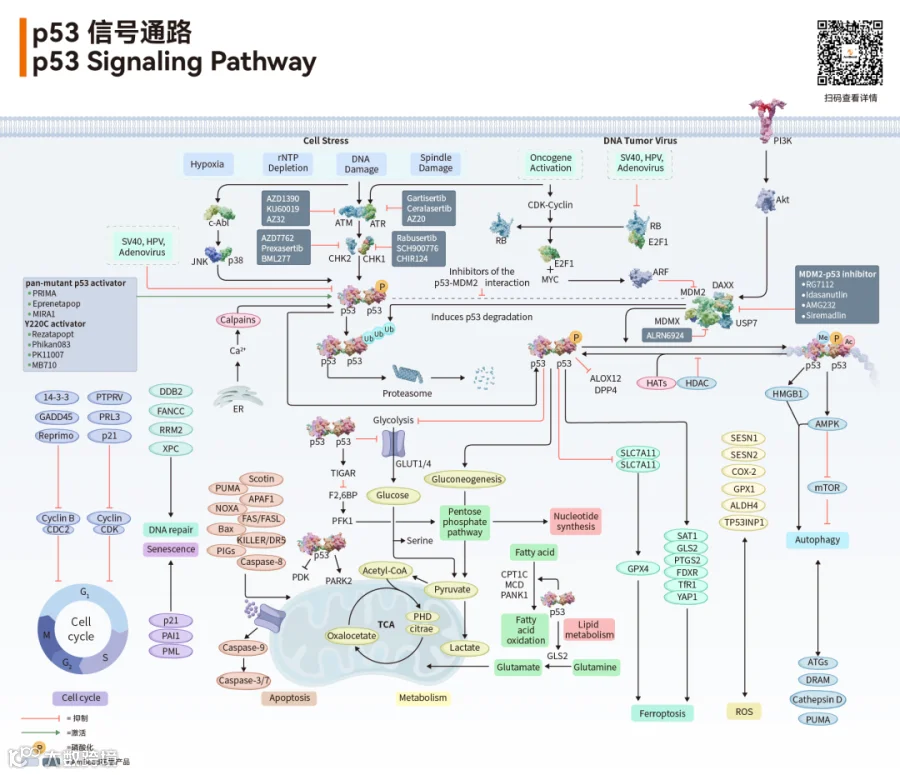
p53信号通路的稳态依赖于精密的负反馈调控环路:静息状态下,E3泛素连接酶MDM2与p53结合并促进其降解;同时MDM2是p53的转录靶基因,p53激活后诱导MDM2表达,构成经典的负反馈调节环路。MDMX作为MDM2的同源蛋白,通过形成异源二聚体增强对p53的抑制[2]
多种应激信号可解除MDM2/MDMX的抑制,激活p53:DNA损伤通过ATM/ATR激酶磷酸化p53(Ser15/20)和MDM2,阻断二者结合;癌基因激活通过ARF蛋白隔离MDM2;核糖体应激通过核糖体蛋白L5/L11/L23抑制MDM2功能[2]
2.下游效应网络
p53作为转录因子,直接调控超过100个靶基因的表达,广泛参与细胞命运决策[1][2]
基因组稳定性维持:DNA损伤时,p53激活DDB2、XPC等DNA修复蛋白的表达,并诱导p21表达引发细胞周期阻滞,为DNA修复争取时间。
细胞凋亡诱导:当DNA损伤不可修复时,p53通过转录激活PUMA、BAX、NOXA等促凋亡基因,启动线粒体凋亡通路。
细胞衰老:p53介导的永久性细胞周期停滞是重要的肿瘤抑制机制之一。
铁死亡调控:p53通过抑制SLC7A11表达减少胱氨酸摄取,进而抑制GPX4活性,促进脂质过氧化和铁死亡发生[3][5]
代谢重编程:p53通过抑制糖酵解相关基因表达,促进氧化磷酸化和线粒体功能,同时调控脂肪酸合成和胆固醇代谢[2][4]
肿瘤免疫微环境调控:p53通过调节细胞因子表达、抗原提呈和免疫检查点分子,影响肿瘤免疫应答[2][5]
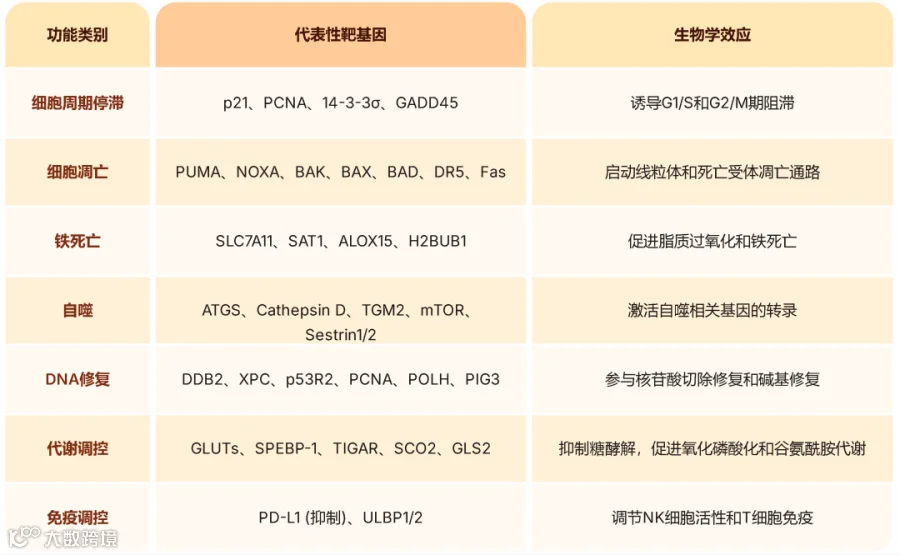
图3. p53下游效应网络代表性靶基因及功能[1][2]
Section.03
p53核心进展与研究热点
1.靶向突变p53再激活:首个选择性药物进入II期临床
针对Y220C热点突变(在p53表面产生疏水空腔),首个口服选择性再激活剂Rezatapopt(PC14586) 取得里程碑进展。II期PYNNACLE试验在所有队列的97例可评估患者中,观察到客观缓解率 (ORR)为33% ,中位缓解持续时间为6.2个月。其中卵巢癌亚组ORR为43%,中位缓解持续7.6个月[6]。此外,三氧化二砷(ATO) 被证实可结合p53核心结构域上的隐蔽别构位点,稳定多种结构型突变(如R175H、G245S、R249S),恢复其转录功能,已进入临床试验评估[2][7]。APR-246(Eprenetapopt) 作为广谱突变再激活剂,与阿扎胞苷联合治疗TP53突变MDS/AML患者显示出良好疗效,但其III期试验因缺乏分层而未达主要终点,提示精准分层的迫切性[2][8]。
2.MDM2抑制剂的优化与PROTAC策略
对于保留野生型p53的肿瘤,RG7388(idasanutlin) 等第二代MDM2抑制剂通过模拟p53的Phe19、Trp23、Leu26三个残基插入MDM2疏水裂隙,在临床前模型中显示高效抗肿瘤活性。然而,III期MIRROS试验中RG7388联合阿糖胞苷未显著改善R/R AML患者的总生存期(8.3 vs 9.1个月)[1][2]。AMG-232和APG-115等其他MDM2抑制剂仍在临床探索中。针对长期用药耐药问题,PROTAC技术被用于设计MDM2降解剂(如MD-224),实现p53双重激活[1]。此外,ALRN-6924作为同时靶向MDM2和MDMX的订书肽,在I期试验中展现出可控的安全性和45%的疾病控制率[2]
3.p53在非凋亡性细胞死亡中的调控机制深化
p53的功能已从经典的“基因组守护者”扩展至铁死亡、坏死性凋亡、焦亡等多种非凋亡性细胞死亡的调控[2]
铁死亡是近年研究最为深入的p53调控领域。 p53主要通过抑制SLC7A11表达,减少谷胱甘肽合成,进而抑制GPX4活性,导致脂质过氧化和铁死亡[3]。近期研究表明,中华猕猴桃根提取物通过上调p53并下调SLC7A11/GPX4轴诱导结直肠癌细胞铁死亡[9]。同期研究发现,褐藻来源的岩藻黄质通过激活p53/SLC7A11/GPX4轴在咽下鳞癌细胞中诱导铁死亡[10]。这些研究为开发基于p53-铁死亡轴的天然产物抗癌药物提供了实验依据。
坏死性凋亡方面,p53可通过上调PUMA、TLR3或长链非编码RNA NRF间接激活RIPK1/RIPK3-MLKL通路[5]。在心肌缺血再灌注损伤和急性胰腺炎模型中,p53依赖的坏死性凋亡是关键病理机制之一。焦亡方面,p53可转录激活NLRP3、Caspase-1和GSDME等焦亡执行分子[5]。在结直肠癌中,人参皂苷Rh3通过激活p53-NRF2通路同时诱导焦亡和铁死亡,协同抑制肿瘤。
4.p53靶向免疫治疗:从新抗原到TCR-T
p53在肿瘤免疫中的角色日益凸显[5]。TP53突变产生的共享新抗原可作为“公共新抗原”开发“货架式”免疫疗法,覆盖不同患者群体。热点突变R175H、Y220C、R273H和G245S衍生的p53肽段在多种肿瘤类型中均有表达,使其成为极具吸引力的免疫治疗靶点。基于mRNA的治疗性疫苗和TCR工程化T细胞技术已将p53靶向免疫治疗从概念推向临床相关策略。尽管p53靶向免疫治疗在早期临床试验中已显示出安全性和诱导抗原特异性T细胞应答的能力,但客观肿瘤反应仍较为有限,亟需进一步优化。
📚 参考文献
[1] Signal Transduct Target Ther. 2023 Mar 1;8(1):92.
[2] Signal Transduct Target Ther. 2026 Apr 6;11(1):121.
[3] Nature. 2015 Apr 2;520(7545):57-62.
[4] Semin Cancer Biol. 2022 Oct;85:4-32.
[5] Nat Rev Mol Cell Biol. 2025 Aug;26(8):600-614.
[6] News release. PMV Pharmaceuticals. Published September 10, 2025.
[7] Nat Rev Cancer. 2021 Mar;21(3):141.
[8] J Clin Oncol. 2021 May 10;39(14):1584-1594.
[9] Front Pharmacol. 2026 Jan 14;17:1724983.
[10] Mar Drugs. 2026 Jan 27;24(2):55.
| AmBeed产品推荐 |
|
|
|
|






往期推荐
不止于经典:解密cAMP信号,洞悉其在癌症中的多重调控与治疗新靶点
宫颈癌的“幕后黑手”:HPV是如何一步步“策反”我们身体的?