
第三章 结果

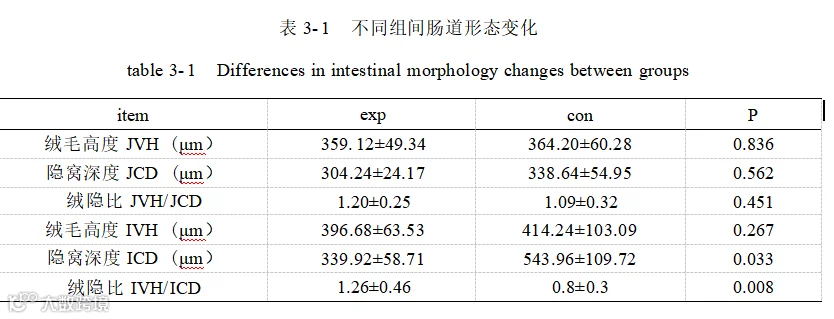
如表 3- 1 所示,酿酒酵母添加组(exp)和基础日粮组(con)回肠隐窝深度显著降低(P<0.05),回肠绒隐比极显著升高(P<0.01)。空肠隐窝深度、空肠绒毛高度、空肠绒隐比、回肠绒毛高度数据均无明显差异(P>0.05)。
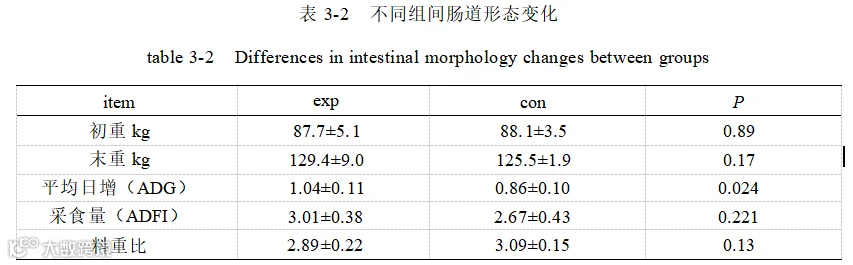
如表 3-2 所示,添加酿酒酵母组(exp)和基础日粮组(con)、平均日增重(ADG)显著升高(P <0.05),结测体重、平均日采食量(ADFI)有所升高,料重比降低但均无明显差异(P >0.05)。
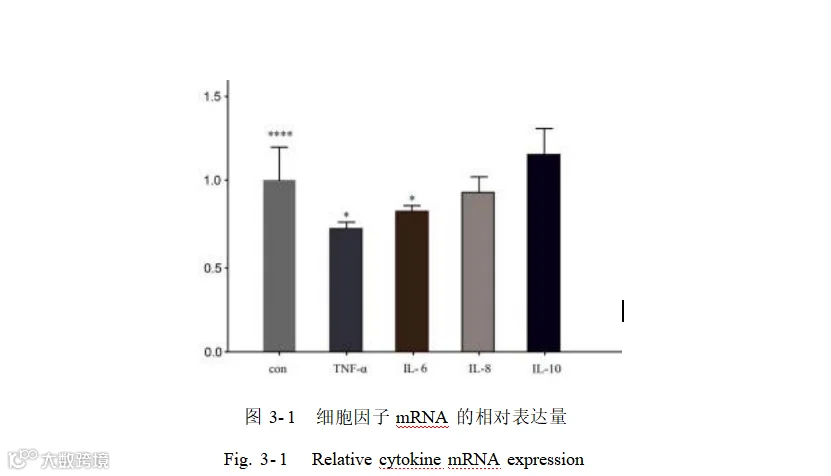
酿酒酵母添加组中白细胞介素-6(IL-6)、肿瘤坏死因子-α(TNF-α) 的 mRNA相对表达量有显著降低(P<0.05)。白细胞介素-1β(IL- 1β) 虽有下降但下降不显著,白细胞介素-10(IL- 10)有上升但差异不显著(P>0.05)。图 3- 1
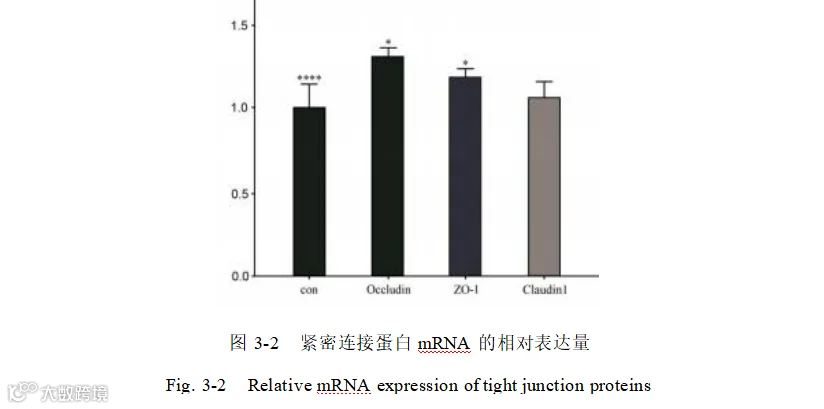
我们研究了酿酒酵母添加组紧密连接蛋白相关基因的相对表达量其中添加组的Occludin和ZO- 1的mRNA的相对表达量显著升高(P<0.05)。Claudin-1的 mRNA的相对表达量虽升高但不显著(P>0.05)。图 3-2
对 10 个育肥猪盲肠内容物样本进行 Illumina HiSeq 测序(MGI DNBSEQ-T7 平台进行测序),下机后共得到 828 ,613 ,688 条序列,平均每个样本 82 ,861 ,369 条序列。经过质量控制及去除宿主污染后,共得到 776,922,494 条高质量序列,平均每个样本 77 ,692 ,249 条序列,数据拼接组装后,共得到 7 ,092 ,972 个 contigs ,平均每个样本 709 ,297 个,其中 N50 平均为 1 ,488。
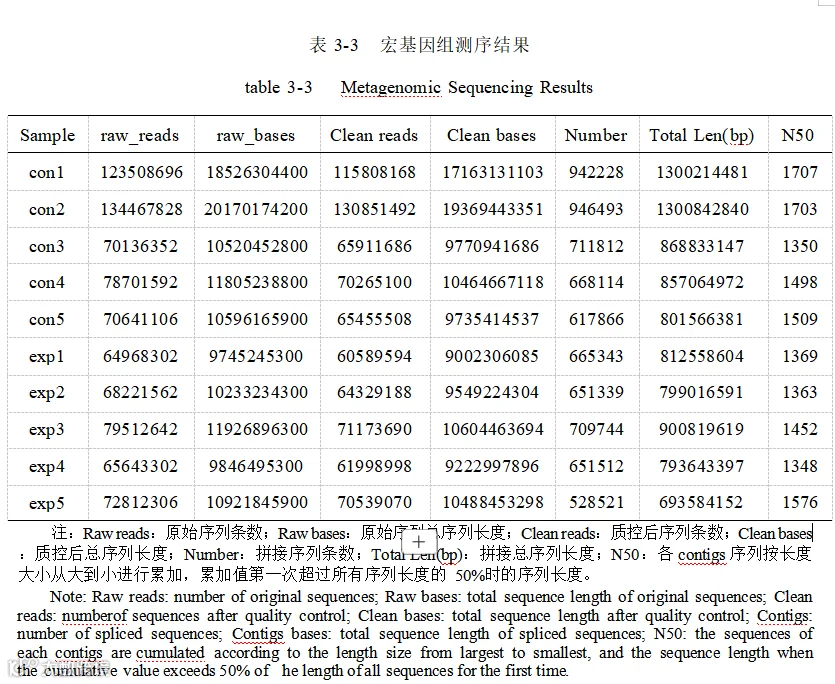
通过 NR 数据库获得物种注释,结果见图 3-3 显示,基础日粮组猪盲肠微生物中细菌相对丰度最高,平均占总微生物相对丰度的 83.29% ,病毒、古细菌及真核生物分别为 14.81% 、1.6%和 0.3% ,酿酒酵母添加组中细菌、病毒、古细菌及真核生物四种物种相对丰度分别是 87.04% 、11.86% 、0.96% 、0. 14% 。加酿酒酵母组和基础日粮组一共检测到种(Species)水平共 31713 个,卡方检测得到添加组和对照组微生物有极显著差异(P<0.001),效应量为中等效应。加酿酒酵母组细菌极显著升高,病毒极显著降低。
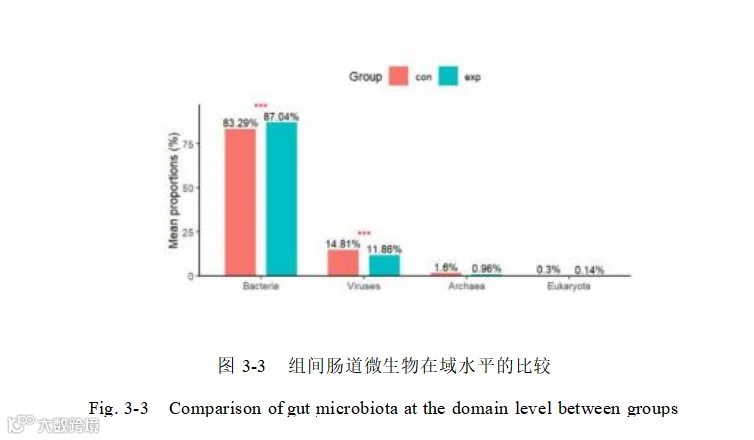
对宏基因组结果中的细菌进行分析,共鉴定出 162 个门,205 个纲,303 个目, 758 个科,3 ,589 个属,25859 个菌种,结果表明对添加酿酒酵母组和基础日粮组细菌组成模式明显不同。在门水平上(图 3-4 左),Bacillota(厚壁菌门)、Bacteroidota (拟杆菌门)、Spirochaetota(螺旋体门)、Pseudomonadota(假单胞菌门),是两组中的优势菌门,添加酿酒酵母组和基础日粮组的 Bacillota(厚壁菌门)丰度最高分别是 51.66%和 55.03% ,其次是 Bacteroidota(拟杆菌门)添加组高于对照组分别是37.20%和 33.01% ,两组细菌未达到统计学显著性(p=0.089>0.05),但接近临界值,可能因样本量不足或效应较弱。
在属水平上(图 3-4 右)优势属具有明显的差异。Prevotella(普雷沃氏菌属)、 Clostridium(梭菌属)、Bacteroides(拟杆菌属)、Treponema(密螺旋体属)、Segatella为添加酿酒酵母组和基础日粮组的优势菌属,其中优势菌属 Segatella在对照组中仅有 2.08%而加酿酒酵母组该菌属比例为 8.67% ,极显著高于对照组(P<0.01)。添加组中 Segatella、Palleniella 等 21 个菌属显著高于对照组(P<0.05),Bullifex、Eubacterium等 18 个差异菌属显著低于对照组(P<0.05)。
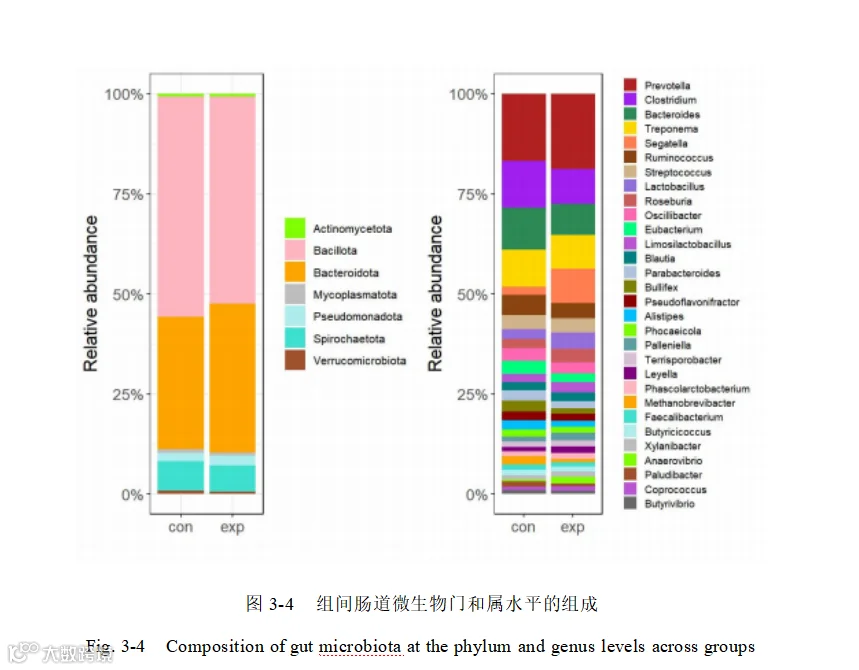
用基于 Bray-Curtis 距离的 ANOSIM(Analysis of similarities)相似性分析进一步验证表明,添加酿酒酵母组和基础日粮组之间物种(R= 0.184,P= 0.046)(图 3-5 左)及基因(R= 0.26 ,P = 0.008)相对丰度差异显著大于组内差异(图 3-5 右)。
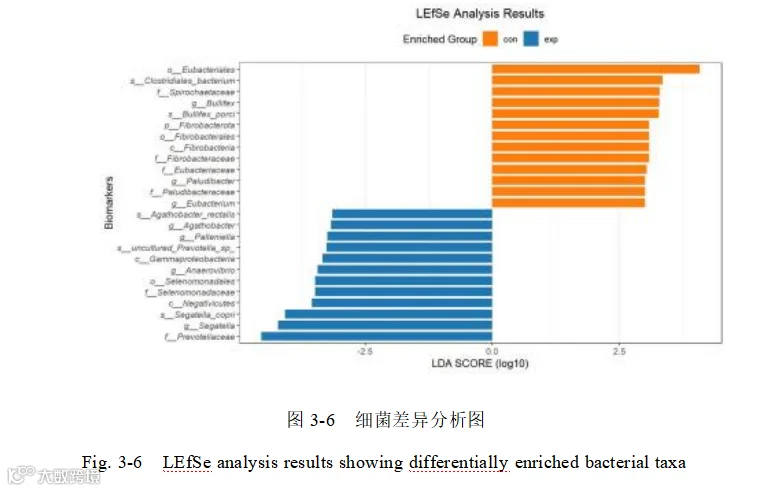
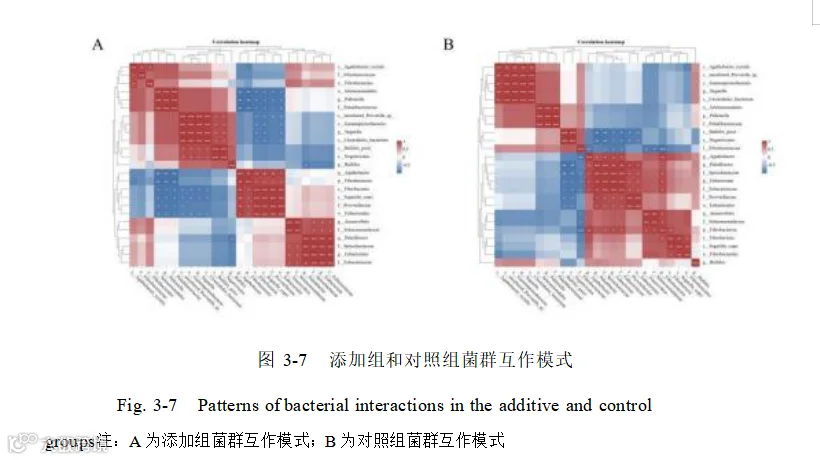
使用“核心微生物群 ”相对丰度前 1000的物种进行 LEfSe 分析(LDA> 2.0),鉴定出了 140 个差异的细菌种类。选择 LDA> 3.0,mean>1000 共 25 个高差异的细菌种类做 LDA 差异分析图(图 3-6),再对 25 个差异菌种进行相关性分析,了解添加酿酒酵母组和基础日粮组组内菌群相互作用的方式。(图 3-7)
LEfSe 菌属差异分析添加酿酒酵母组的差异指示菌属是普雷沃菌属(Segatella)、厌氧弧菌属(Anaerovibrio) 、阿加托巴克特属(Agathobacter) 、Palleniella;差异指示种水平物种是普雷沃菌种(Segatella copri)、阿加托巴克特直肠菌(Agathobacter rectalis)和 Prevotella sp 。基础日粮组差异指示菌属是沼泽杆菌属(Paludibacter) 、欧文氏菌属 (Eubacterium)和 bulllifex;差异指示种水平物种是梭菌目细菌(Clostridiales bacterium)和 bulllifexporci。
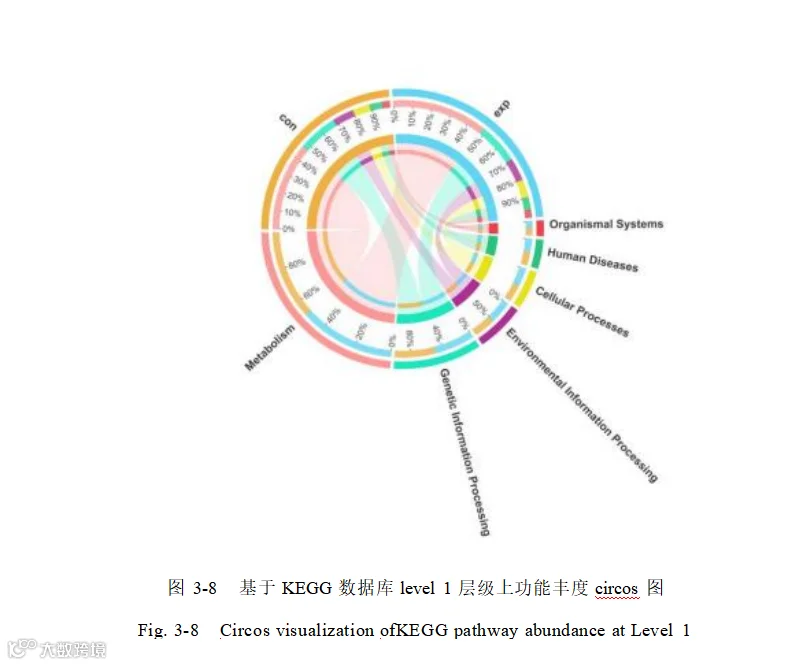
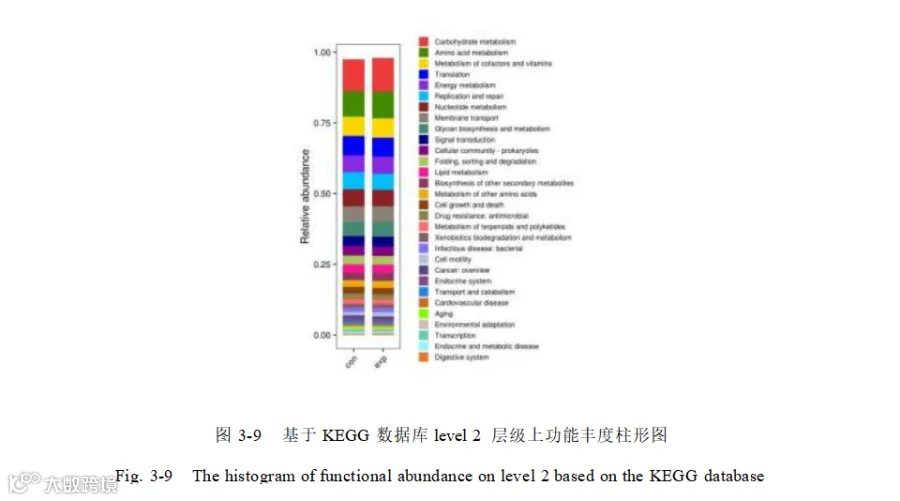
对两组猪的盲肠微生物功能进行了KEGG 功能注释,在 KEGG Level 1 通路中路中(图 3-8),新陈代谢(Metabolism ,47.94%)在各日龄中平均相对丰度最高,其次 为 遗传信 息 处 理( Genetic information processing ,21.0% ) 、 环 境信 息 处 理(Environmental information processing,11.44%)、细胞过程(Cellular processes,9. 10%)、人类疾病(Human diseases,6.85%)以及有机体系统(Organismal systems,3.67%)。在二级通路中(图 3-9),共注释到 47个 KEGG Level 2 通路,平均相对丰度前五的代谢通路为氨基酸代谢(Amino acid metabolism ,10.46%) 、碳水化合物代谢(Carbohydrate metabolism,10.37%)、辅因子和维生素代谢(Metabolism of cofactors and vitamins,6.83%)、翻译(Tanslation,6.73%)以及能量代谢(Energy metabolism , 6.03%)。
对所有level 1 功能进行了 LEfSe 分析(LDA> 2.0),鉴定出 2 个相关的细菌功能,添加组注释到的是代谢(Metabolism)对照组是人类疾病(Human Diseases)。
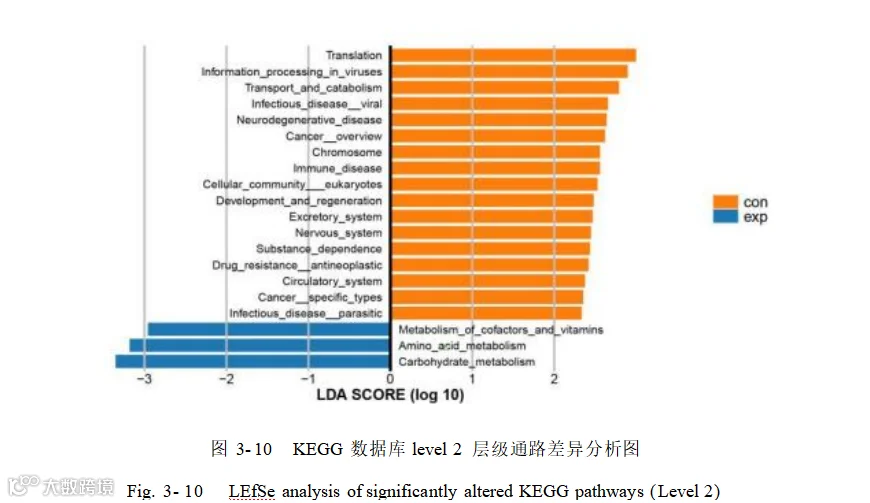
对所有 level 2 功能进行了 LEfSe 分析(LDA> 2.0),鉴定出 20 个相关的细菌功能(图 3- 10) ,其中添加组关注到的通路为辅因子和维生素代谢(Metabolism of cofactors and vitamins)、氨基酸代谢(Amino acid metabolism)、碳水化合物代谢(Carbohydrate metabolism)。对照组关注到的通路为翻译(Translation)、病毒中的信息处理( Information processing in viruses ) 、运输和分解代谢(Transport and catabolism ) 、 病 毒 性 传 染 病 ( Infectious disease: viral ) 、 神 经 退 行 性 疾 病(Neurodegenerative disease)、癌症概述(Cancer: overview)、染色体(Chromosome)、免疫疾病(Immune disease)、细胞群体–真核生物(Cellular community –eukaryotes)、发育与再生(Development and regeneration)、排泄系统(Excretory system)、神经系统(Nervous system)、物质依赖(Substance dependence)、抗肿瘤药物耐药性(Drug resistance: antineoplastic)、循环系统(Circulatory system)、特定类型癌症(Cancer: specific types)以及寄生虫性传染病(Infectious disease: parasitic)。
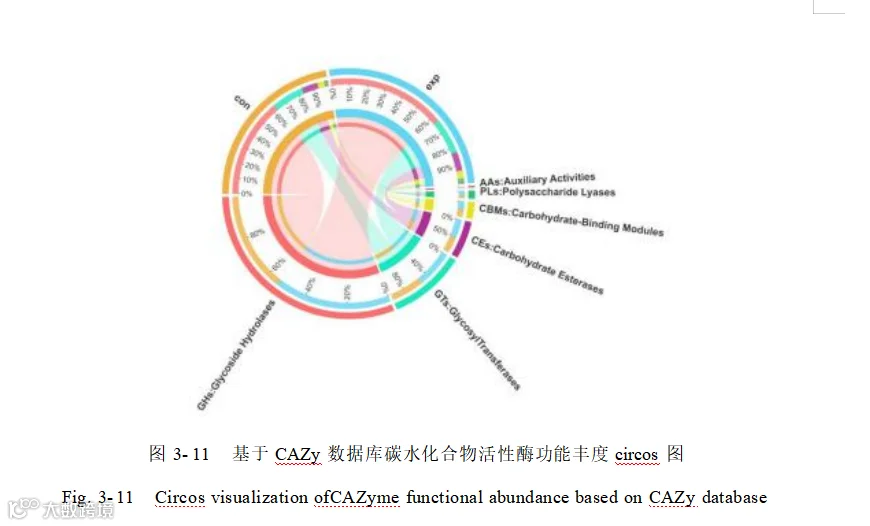
将基因与碳水化合物活性酶数据库 CAZy 进行比对,碳水化合物活性酶注释结果显示(图 3- 11),各日龄中的糖苷水解酶(GHs ,63.09%)相对丰度最高,其次是糖基转移酶(GTs,20.31%)、碳水化合物酶(CEs,9.94%)、碳水化合物结合模块(CBMs, 4.40%)、多糖裂解酶(PLs ,2.02%)以及 辅助活性家族(AAs ,0.24%)。分别在对照和添加组中鉴定到 424 、与 422 种碳水化合物酶,特异性酶分别为 3 和 1 种,共有酶为 421 种。添加组中特有CBM46 、GH5_ 18 、PL22_2 对照组中特有 GH13_ 13。
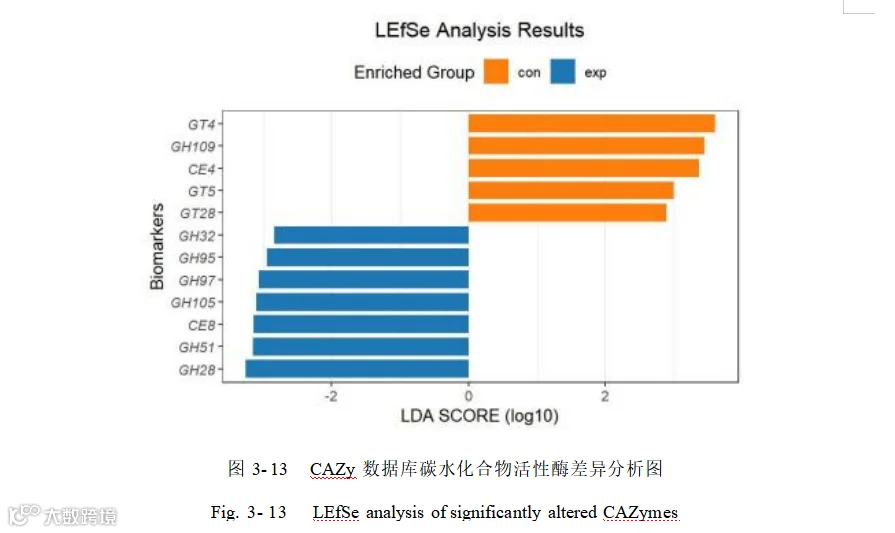
将对照组和添加组中相对丰度前 30 的碳水化合物酶进行差异比较,并用热图展示(图 3- 12)。相对丰度前 30 的碳水化合物酶进行 LEfSe 分析(LDA> 2.0 图 3- 13)相对丰度前 30 中有 12 个碳水化合物酶的相对丰度在不同组中差异显著(P < 0.05),多数碳水化合物酶糖苷水解酶(GHs)在添加组丰度较高其中 GH97 、GH28 、GH51、GH32 、GH95 、GH105和CEs 的 CE8 富集在添加酿酒酵母组。糖基转移酶(GTs)中GT4 、GT28 、GT5 ,CEs 中 CE4 和 GHs 中 GH109 富集在对照组中。

将预测基因的蛋白序列比对到 CARD-v3.1.4 库,抗性基因注释结果显示(图 3- 14),共检测到 39 种抗性基因抗生素,其中检测到的抗生素抗性基因丰度最高的为四环素类抗生素(Tetracycline antibiotic ,12.66%),其次为大环内酯类抗生素(Macrolide antibiotic ,10.62%)、氟喹诺酮类抗生素(Fluoroquinolone antibiotic ,7.98%)、青霉烷类(Penam ,5.99%)以及肽类抗生素(Peptide antibiotic ,5.78%)等。
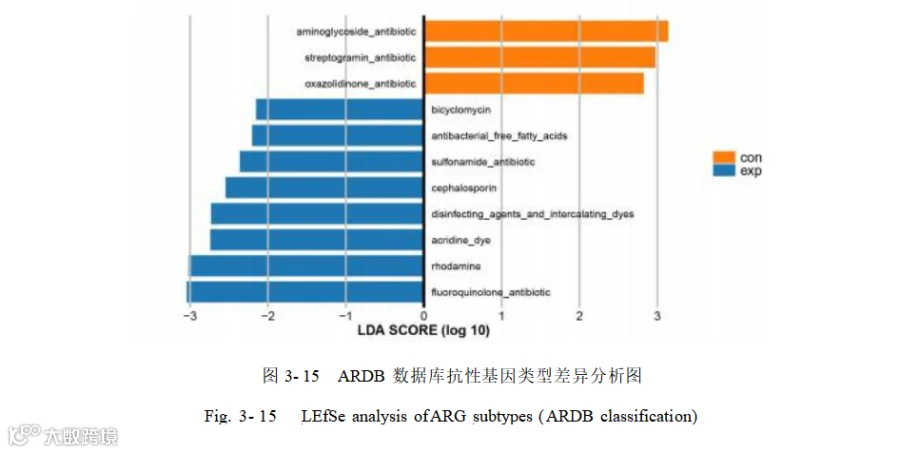
对 39种抗性基因抗生素进行了 LEfSe 分析(LDA> 2.0),鉴定出 11 个显著性差异的抗生素抗性基因(图 3- 15),其中富集到添加组的抗性基因有:氟喹诺酮类抗生素(Fluoroquinolone antibiotic ) 、头孢菌素(Cephalosporin ) 、磺胺类抗生素(Sulfonamide antibiotic)、罗丹明(Rhodamine)、吖啶染料(Acridine dye)、消毒剂 与 嵌 入 染 料(Disinfecting agents and intercalating dyes ) 、 抗 菌 游 离 脂 肪 酸(Antibacterial free fatty acids)、双环霉素(Bicyclomycin)。富集到对照组的有氨基糖苷类抗生素(Aminoglycoside antibiotic)、链霉素类抗生素(Streptogramin antibiotic)、恶唑烷酮类抗生素(Oxazolidinone antibiotic)。
分别在分别在对照和添加组中鉴定到 1057 与 1046 种抗性基因类型,特异性抗性基因类型分别为 29 与 18 种,共有抗性基因类型为 1028 种(图 3- 16)。其中,ALI- 1、rmtG和vatA等 29 个抗性基因为对照组中特有,CARB-5 、MOX-5 和 fusC 等 18 个抗性基因为添加组所特有。筛选 P<0.05 的抗性基因共 42 个,其中 bcrA 、baeS 等在对照组中丰度较高,arlS 、optrA 等在添加组中丰度较高。


非靶向代谢组
对 QC 样本进行皮尔逊(Pearson)相关性分析,QC 样本相关性越高(| r |越接近于 1)说明整个检测过程稳定性越好,数据质量越高(图 3- 17)。
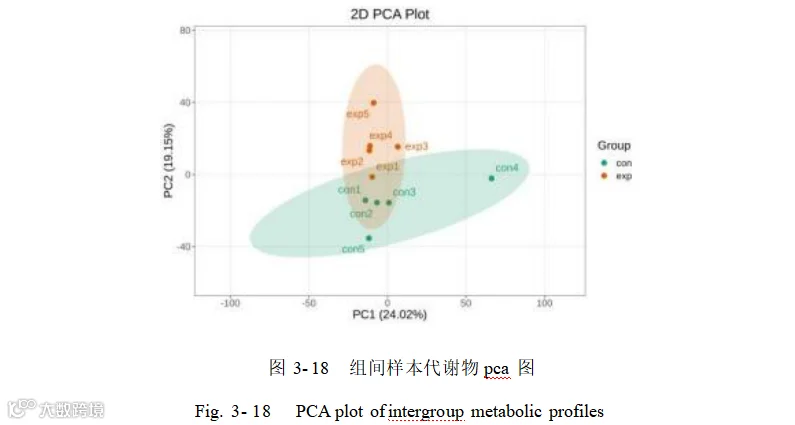
对添加组和对照组的盲肠内容物的代谢物进行主成分分析,主成分分析结果显示(图 3- 18)
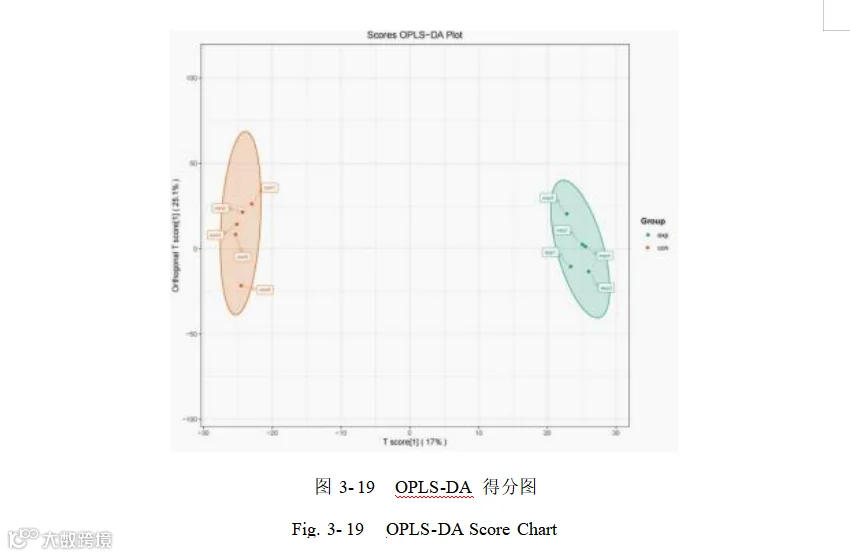
通过构建有监督的正交偏最小二乘法判别分析模型,来进一步比较添加酿酒酵母组和基础日粮组组间代谢物差异。OPLS-DA 得分图中的横坐标表示预测成分得分值,可以看出添加酿酒酵母组和基础日粮组组间的差距;纵坐标表示正交成分得分值,可以看出添加酿酒酵母组和基础日粮组各组内的差距。OPLS-DA 模型显示,添加酿酒酵母组和基础日粮组组间存在差异(图 3- 19)。
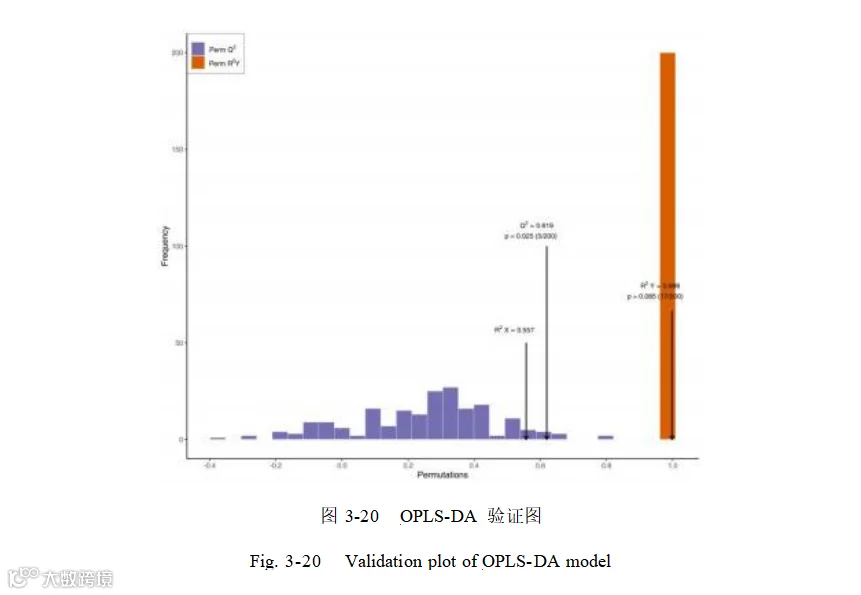
正负离子模式下,Q²为 0.619,Q²大于 0.5,提示正负离子模式下添加组和对照组OPLS-DA 模型为有效模型(图 3-20)。
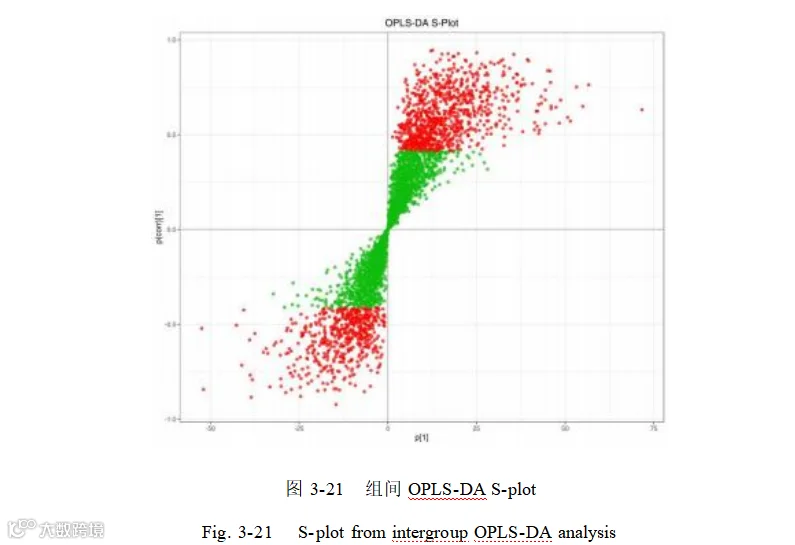
使用 S-plot 图展示 OPLS-DA 模型下,横坐标表示主成分与代谢物的协方差,纵坐标表示主成分与代谢物的相关系数,越靠近右上角和左下角的代谢物表示其差异越显著。S-plot 图结果显示,正负离子模式下产仔数高低组间代谢物存在差异,红色的点表明这些代谢物的VIP值大于1,绿色的点表示这些代谢物的VIP值小于等于 1 (图 3-21)
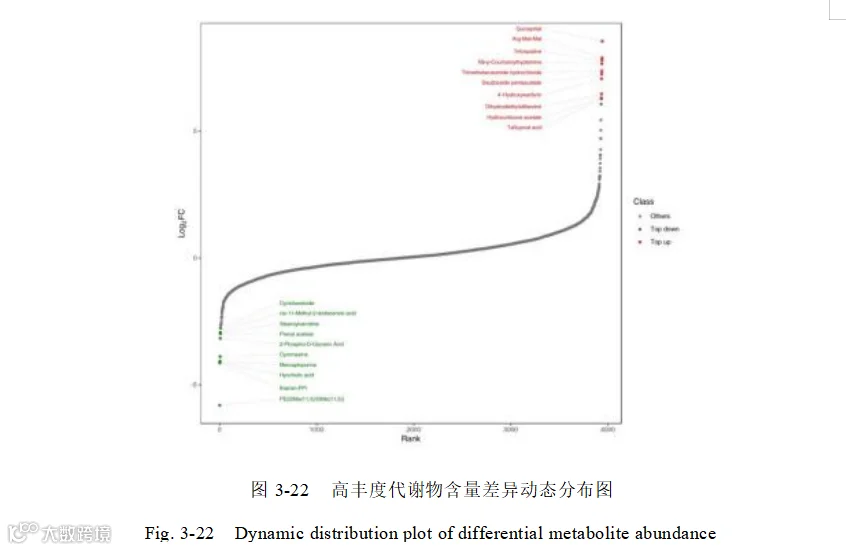
上调的高丰度差异代谢物主要包括:他氟前列素酸(Tafluprost acid),皮质醇21-乙酸酯(Hydrocortisone acetate),二氢己烯雌酚(Dihydrodiethylstilbestrol),4-羟基-3-[1-(4-羟基苯基)-3-氧代丁基]-2H-铬-2-酮(4'-Hydroxywarfarin),五乙酸德乌齐糖苷(Deutzioside pentaacetate) ,Tribenzagan Hydrochloride(Trimethobenzamide hydrochloride),N-阿魏酰色胺(Nb-p-Coumaroyltryptamine),曲喹啉(Tritoqualine), L-精氨酰-L-甲硫氨酰-L-甲硫氨酸(Arg-Met-Met),喹那普利拉(Quinaprilat)。
下调的高丰度差异代谢物主要包括:PE(DiMe(11,5)/DiMe(11,5)) ,硫胺素-PPi (thiamin-PPi),球胆酸(Hyocholic acid),6-巯基嘌呤;巯基嘌呤(Mercaptopurine),灭蝇胺(Cyromazine),2-磷酸-D-甘油酸(2-Phospho-D-Glyceric Acid),醋酸异戊二烯(Prenyl acetate),硬脂酰肉碱(Stearoylcarnitine),顺式- Δ2- 11-甲基十二碳烯酸(cis-11-Methyl-2-dodecenoic acid),放线菌酮(Cycloheximide)。代谢物含量差异动态分布图 3-22。
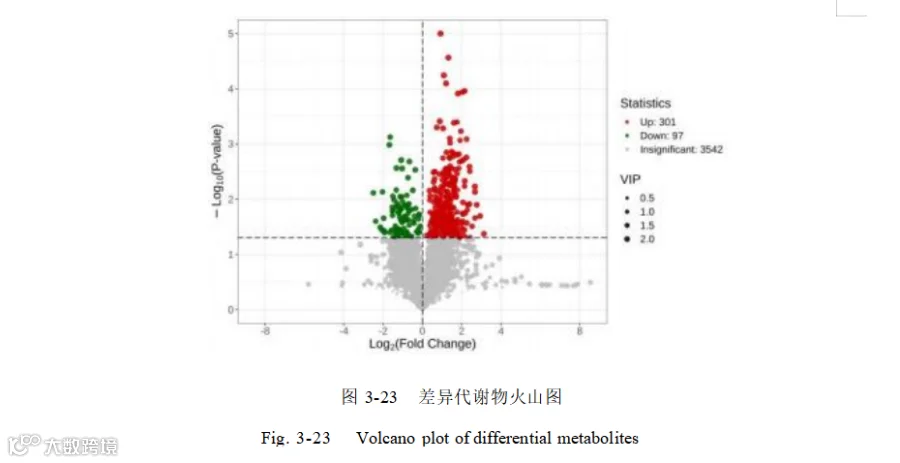
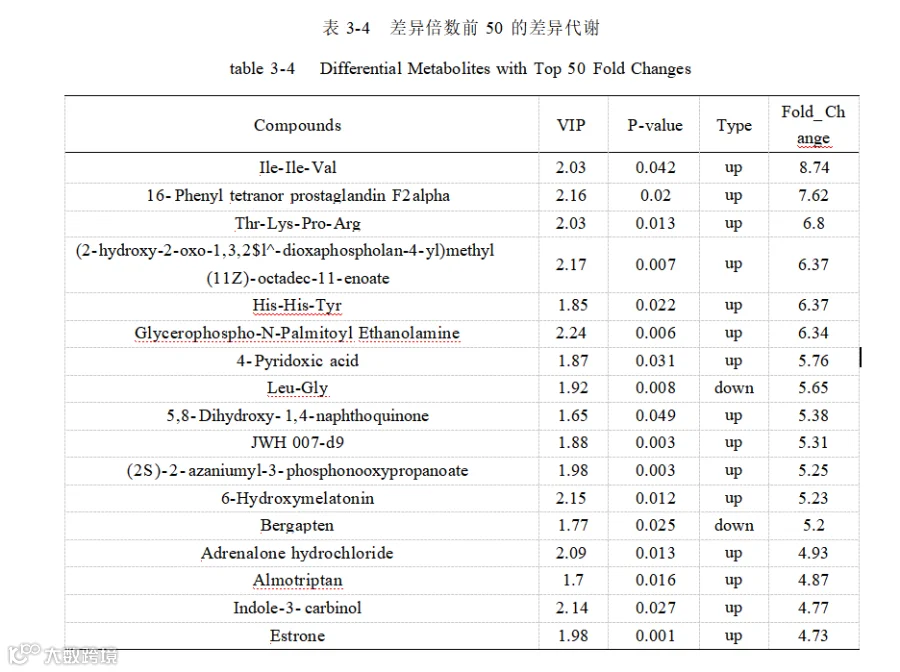
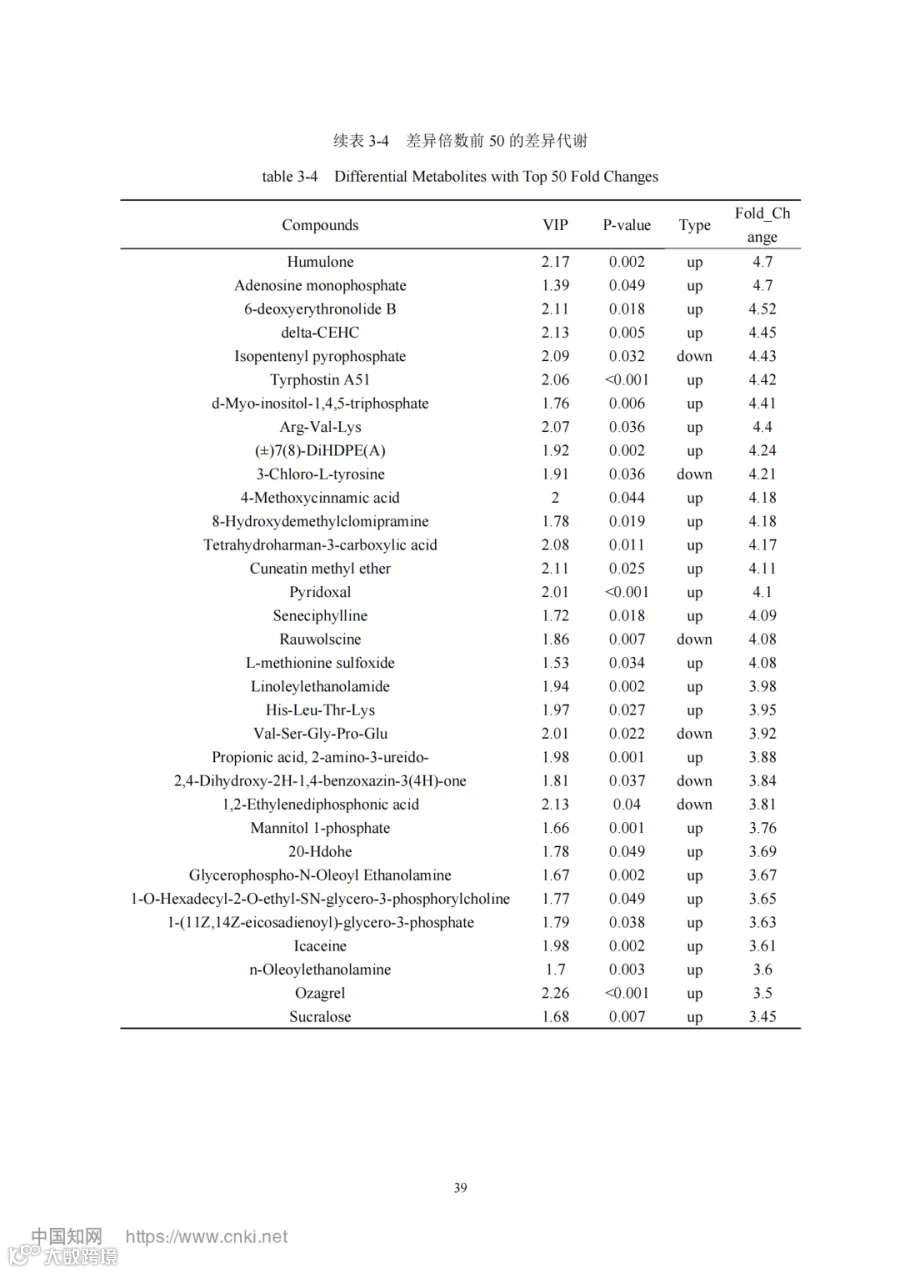
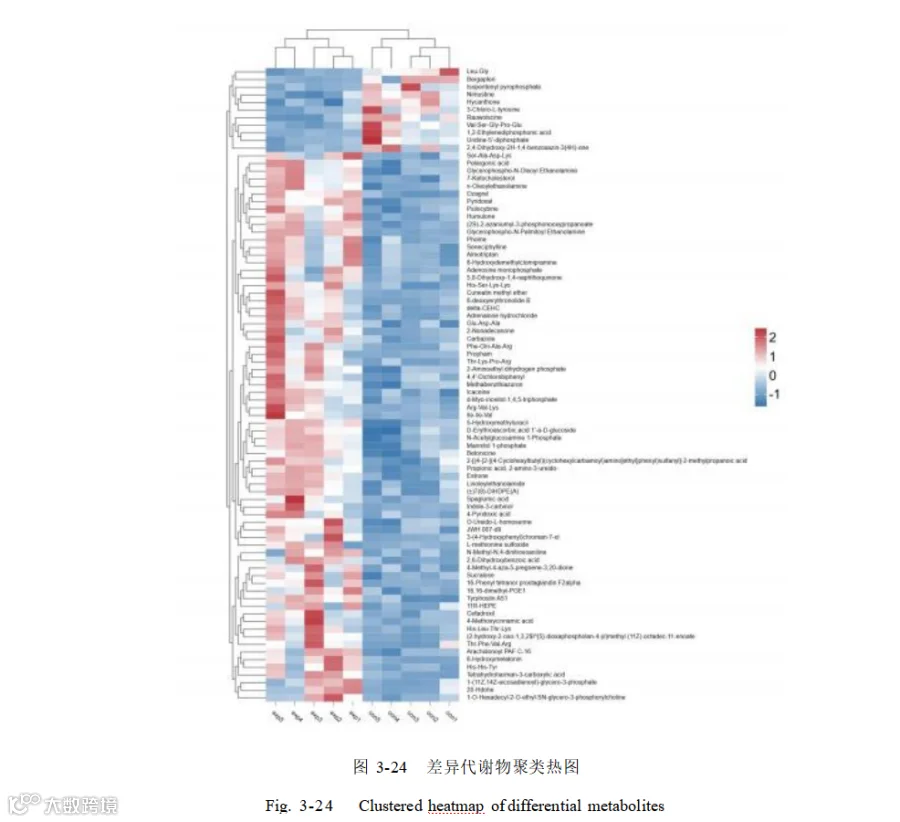
添加酿酒酵母组和基础日粮组共筛选出代谢物 3940 个,基于 OPLS-DA 模型,得到的变量重要性投影(VIP)选取 VIP > 1 、P<0.05 的代谢物为显著差异代谢物,共筛选出 398个差异代谢物,其中上调的有 301 个,下调的有 97 个如图 3-23。其中上调的代谢物主要有:L-异亮氨酸-L-异亮氨酸-L-缬氨酸(Ile-Ile-Val)、16-苯基四去甲 前 列 腺 素 F2α ( 16-Phenyl tetranor prostaglandin F2alpha ) 、 Thr-Lys-Pro-Arg 、 (11Z)-octadec-11-enoate 、组氨酸-组氨酸-酪氨酸(His-His-Tyr)、甘油磷酸-N-棕榈酰乙醇胺(Glycerophospho-N-Palmitoyl Ethanolamine)、4-吡哆酸(4-Pyridoxic acid)、 5,8-二羟基-1,4-萘醌(5,8-Dihydroxy- 1,4-naphthoquinone)等。下调的主要有 L-亮氨酰-L- 甘氨酸(Leu-Gly ) ,佛手柑 内酯(Bergapten )异戊烯焦磷酸(Isopentenyl pyrophosphate),3-氯-L-酪氨酸(3-Chloro-L-tyrosine) 。表 3-4 展示了差异倍数前50 的差异代谢物。
以添加组 vs 对照组 Fold_Change>3 和 Fold_Change<0.333、VIP > 1 且 P <0.05 为标准筛选代谢物做热图(图 3-24),其中代谢物亮氨酰甘氨酸(Leu-Gly)、佛手柑内酯(Bergapten)、异戊烯基焦磷酸(Isopentenyl pyrophosphate)、尼莫司汀(Nimustine)、海坎酮(Hycanthone)、3-氯-L-酪氨酸(3-Chloro-L-tyrosine)、萝芙木碱(Rauwolscine)、缬氨酰丝氨酰甘氨酰脯氨酰谷氨酸(Val-Ser-Gly-Pro-Glu ) 、 1,2- 亚乙基二膦酸(1,2-Ethylenediphosphonic acid)、尿苷-5'-二磷酸(Uridine-5'-diphosphate)、2,4-二羟基-2H- 1,4-苯并噁嗪-3(4H)-酮(2,4-Dihydroxy-2H- 1,4-benzoxazin-3(4H)-one)在对照组中显著富集,其余代谢物富集在添加组。
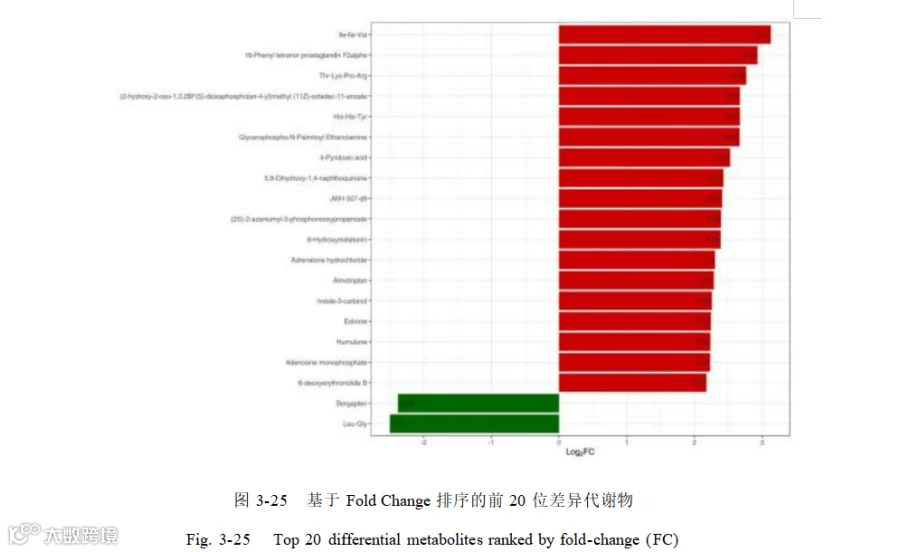
选取差异代谢物中 Fold_Change 排名前 20 作为标志差异代谢物做后续的联合分析。差异倍数排名前 20 的差异代谢物如图 3-25 所示。
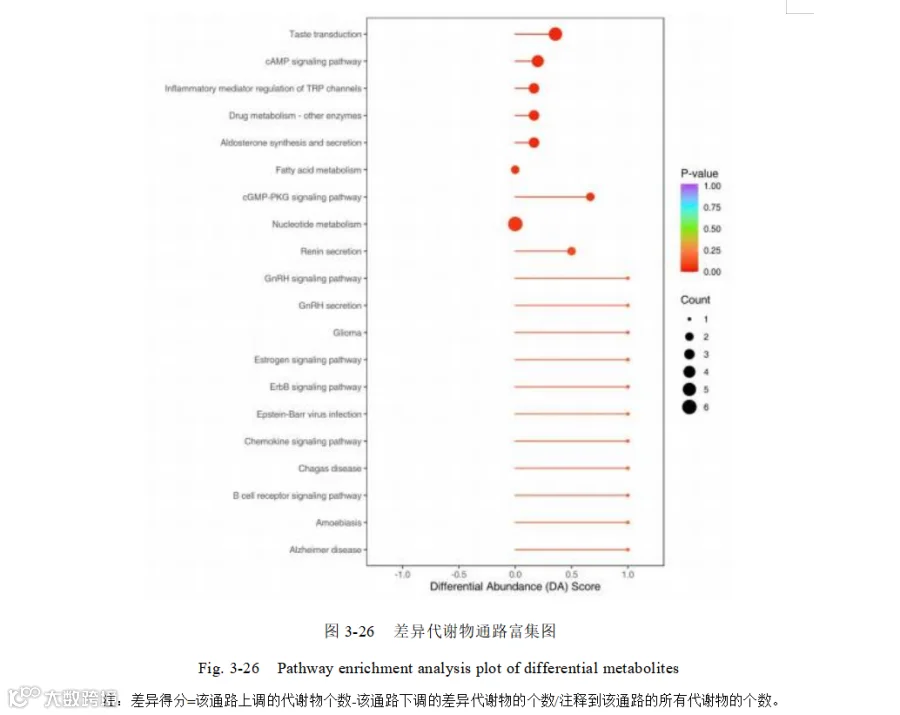
本研究通过对筛选到的差异代谢物进行 KEEG 富集(图 3-26),共富集到 164 条代谢通路。通过差异丰度得分来捕捉该通路所有代谢物的总体变化,结果发现,倾向于上调的差异代谢通路主要有:
味觉转导通路(Taste transduction)、cAMP 信号通路(cAMP signaling pathway)、药物代谢-其他酶(Drug metabolism - other enzymes)、醛固酮合成与分泌(Aldosterone synthesis and secretion)、TRP 通道的炎症介质调控(Inflammatory mediator regulation of TRP channels) 、脂肪酸代谢(Fatty acid metabolism) 、cGMP-PKG 信号通路(cGMP-PKG signaling pathway)、核苷酸代谢(Nucleotide metabolism)、肾素分泌(Renin secretion)、MAPK 信号通路(MAPK signaling pathway)、ErbB 信号通路(ErbB signaling pathway)、Ras 信号通路(Ras signaling pathway)、NF-κB 信号通路(NF-kappa B signaling pathway)、PI3K-Akt 信号通路(PI3K-Akt signaling pathway)、 mTOR 信号通路(mTOR signaling pathway)、中性粒细胞胞外陷阱形成(Neutrophil extracellular trap formation ) 、NOD- 样受体信号通路(NOD-like receptor signaling pathway)、自然杀伤细胞介导的细胞毒性(Natural killer cell mediated cytotoxicity)、 T 细胞受体信号通路(T cell receptor signaling pathway)、Th1 和 Th2 细胞分化(Th1 and Th2 cell differentiation)。
将上述宏基因组标记到的25 种显著差异的菌和代谢组种标记到的20 个差异代谢物质以及表型数据联合分析,横坐标为 25 种差异菌和表型数据,纵坐标为 20 种差异代谢物和表型数据(图 3-27)。
其中回肠隐窝深度 ICD 和普雷沃菌科(Prevotellaceae),真细菌目(Eubacteriales)纤维杆菌门(Fibrobacterota)显著负相关(P<0.05)沼杆菌科(Paludibacteraceae)、 Palleniella 、月牙菌目(Selenomonadales)极显著的正相关(P<0.01)。回肠隐窝深度 ICD 和 6-脱氧赤酮内酯 B(6-deoxyerythronolide B)、lle-lle-Val、雌酮(Estrone)、 JWH 007-d9 、极显著负相关(P<0.01)。吲哚-3-甲醇(Indole-3-carbinol)、肾上腺素酮盐酸盐(Adrenalone hydrochloride)、6-羟基褪黑素(6-Hydroxymelatonin)、(2S)-2-铵基-3- 磷酸氧基丙酸((2S)-2-azaniumyl-3-phosphonooxypropanoate ) 、4- 吡 哆酸(4-Pyridoxic acid ) 、甘 油磷 酸-N- 棕榈 酰 乙 醇胺(Glycerophospho-N-Palmitoyl Ethanolamine)、组氨酰-组氨酰-酪氨酸(His-His-Tyr)、(2-羟基-2-氧代-1,3,2$l^-二氧磷杂环戊烷-4-基)甲基(11Z)-十八碳 - 11-烯酸酯((2-hydroxy-2-oxo-1,3,2$l^-dioxaphospholan-4-yl)methyl (11Z)-octadec-11-enoate)、苏氨酰-赖氨酰-脯氨酰-精氨酸(Thr-Lys-Pro-Arg) 、异亮氨酰-异亮氨酰-缬氨酸(Ile-Ile-Val)、佛手柑内酯(Bergapten)显著负相关(P<0.05)。亮氨酰甘氨酸 Leu-Gly极显著正相关(P<0.01)。
回肠绒隐比(IVH/ICD)和代谢物的显著性和回肠隐窝深(ICD)相反。回肠绒隐比(IVH/ICD)与细菌普雷沃菌科(Prevotellaceae)、真细菌目(Eubacteriales)纤维杆菌门(Fibrobacterota)、阿加托杆菌属(Agathobacter)、Segatella copri 均呈现极显正相关(P<0.01),月牙菌目(Selenomonadales)、Palleniella 呈极显著负相关(P<0.01)与代谢物:6-脱氧赤酮内酯 B(6-deoxyerythronolide B)、lle-lle-Val 、雌酮(Estrone)呈极显著正相关。
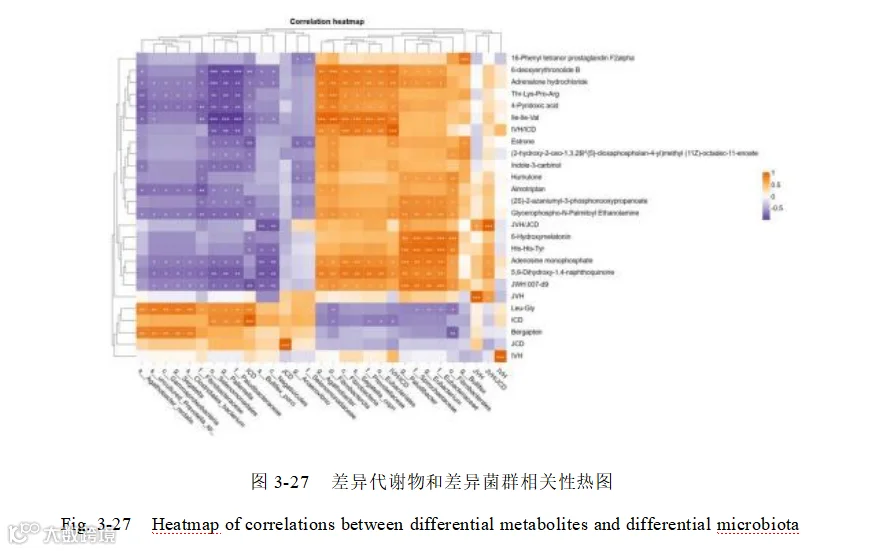
将代谢组中的差异代谢物和宏基因组注释到的差异代谢通路联合分析(图 3-28),辅因子和维生素代谢(Metabolism of cofactors and vitamins)、氨基酸代谢(Amino acid metabolism)和碳水化合物代谢(Carbohydrate metabolism)这三条关键代谢通路均与6-Hydroxymelatonin 和His-His-Tyr显著负相关。
排泄系统(Excretory system)、染色体(Chromosome)、传染病-寄生虫性(Infectious disease - parasitic)、神经系统(Nervous system)、循环系统(Circulatory system)、物质依赖(Substance dependence)、发育与再生(Development and regeneration)、细胞群体-真核生物(Cellular community - eukaryotes)、染色体(Chromosome)、排泄 系 统 ( Excretory system )和代谢物 、 Indole-3-carbinol 、 4-Pyridoxic acid 、 6-deoxyerythronolide B、Adrenalone hydrochloride、Estrone、Thr-Lys-Pro-Arg、lle-lle-Val、 JWH 007-d9 有显著的正相关,他们可能是本试验中关键的枢纽通路和代谢物。
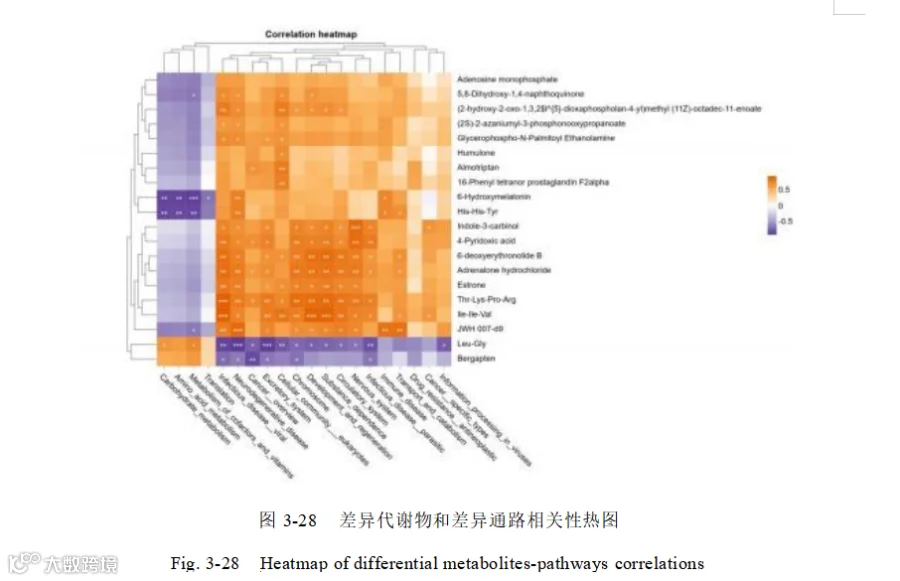
对上述宏基因组筛选到的差异菌群设置条件 LDA>4.0 筛选到的微生物标志物和差异代谢物做相关性网络图 3-29。
Segatella copri 与 60 个代谢物存在显著相关(Spearman 相关系数绝对值大于 0.6, P < 0.05),其中与 16-Phenyl tetranor prostaglandin F2alpha 呈极强的正相关(Spearman相关系数 ρ>0.9 ,P<0.05 ),与2-Aminoethyl dihydrogen phosphate 、Glycerophospho-N-Oleoyl Ethanolamine 、(2S)-2-azaniumyl-3-phosphonooxypropanoate、 5-Hydroxymethyluracil 、Seneciphylline 、8-Hydroxydemethylclomipramine、2,6-Dihydroxybenzoic acid 、N-Methyl-N,4-dinitrosoaniline 、16, 16-dimethyl-PGE1 ,呈强正相关(Spearman 相关系数ρ>0.8 ,P <0.05)。
Segatella 与 26 个代谢物存在显著相关关系,(Spearman 相关系数绝对值大于 0.6,P < 0.05 ),其中与N-Methyl-N,4-dinitrosoaniline 、Glycerophospho-N-Palmitoyl Ethanolamine 具有强负相关(Spearman 相关系数ρ<-0.8 ,P < 0.05)。
普雷沃菌科(Prevotellaceae)和优杆菌目(Eubacteriales)与 24 个代谢物存在显著相关性,(Spearman 相关系数绝对值大于 0.6 ,P < 0.05),其中Seneciphylline 、8-Hydroxydemethylclomipramine、Almotriptan、16-Phenyl tetranor prostaglandin F2alpha,呈强正相关(Spearman 相关系数ρ>0.8 ,P < 0.05)。
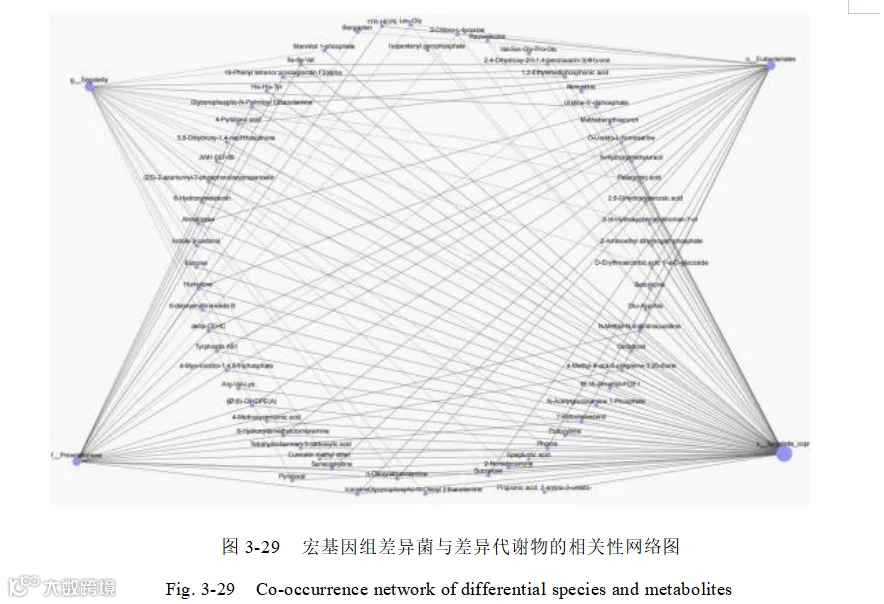
细菌普雷沃菌科(Prevotellaceae)、优杆菌目(Eubacteriales)、Segatella copri、纤维杆菌门(Fibrobacterota)、阿加托杆菌属(Agathobacter)、阿加托巴克特属(Agathobacter)和代谢物 6- 脱氧红霉素B (6-deoxyerythronolide B )、4- 吡 哆酸(4-Pyridoxic acid)、雌酮(Estrone)之间存在共同的、显著正向关联,是影响添加组和对照组之间差异的指示菌和指示代谢物,他们之间的相互作用,共同改善了育肥猪肠道健康,提升了育肥猪的生长性能。
第四章 讨论
酿酒酵母培养物的添加显著增加了平均日增重(ADG)这和梁城的论文观点一致。试验结果显示,酿酒酵母培养物添加组总体采食量上升。可能的原因在于麸皮发酵产物中的活性成分如益生菌、消化酶、以及功能性代谢产物不仅改善了肠道对营养物质的吸收效率,提高了能量和蛋白质的利用率,同时也可能通过调节肠道激素(如GLP- 1 、PYY)的分泌,缩短胃排空时间,减弱饱腹感,从而刺激采食增加。这一观点与赵修报的研究结果相一致。在预试验中,我们观察到添加酿酒酵母会让育肥猪增加摄食次数和进食时长,可能因其特殊气味使得饲料对育肥猪有更大的吸引力。
小肠是吸收营养物质的重要场所,绒毛与微绒毛是小肠的特征结构,其绒毛长度、隐窝深度及绒毛长度/隐窝深度比值能够反映营养吸收效率的高低,其中绒毛长度值越高、隐窝深度值越低、V/C 值越高则表明吸收面积大,对营养物质有更好的吸收。
本试验中的 IVH 无显著性差异,但 IVH/ICD 有极显著性差异,试验结果也表明ICD 有显著的降低。酿酒酵母组 IVH/ICD 回肠绒隐比极显著升高,通常表明肠道吸收功能的优化,提示酿酒酵母培养物可能通过调控隐窝干细胞分化动力学而非单纯促进绒毛伸长来优化肠道功能,隐窝深度的降低反映肠道的改善。隐窝是干细胞增殖的区域,隐窝深度的增加常提示修复损伤或炎症刺激,较浅的隐窝意味着干细胞分化为成熟吸收细胞的速度加快,细胞从隐窝向绒毛顶端迁移的周期缩短,减少未成熟细胞在隐窝中的滞留时间(Peng Li ,2025)。隐窝细胞的增殖是肠道能量的主要消耗部分,该部分代谢能量可以占全身代谢的 20%-30% 。酿酒酵母培养物诱导的隐窝深度降低标志着减少了关于隐窝增殖所需的能量,进而减少了肠道的能量需求。将更多能量分配便可以被直接吸收,促进机体的生长生长,这可能是添加组平均日增重(ADG)显著提升的关键机制(Stoll et al. ,2000)。消化隐窝细胞快速成熟可提高绒毛顶端功能性吸收细胞(如具有丰富转运蛋白的肠上皮细胞)的比例,增强营养吸收效率(陈沁,2021)。尽管绒毛高度未显著增加,但吸收细胞数量的提升可能通过更多的载体介导的主动运输(如葡萄糖、氨基酸的吸收)来增加吸收的效率,再降低料重比。
本研究基于宏基因组测序方法,对添加组和对照组的盲肠内容物进行了系统的研究。我们观察到酿酒酵母添加组和对照组盲肠微生物群落的多样性、结构和功能有显著的差异。与鸟类、兔子及人相似,育肥猪的肠道微生物包括细菌、古菌、真核生物及病毒,其中细菌占主导地位。Lim 等人(2019)分析了从出生至10岁小型猪的肠道微生物群,观察到初生仔猪肠道微生物组多样性随年龄的增大而增大直到约20 周,在之后在整个生命过程中波动较小。本试验选取的是 4 月龄猪,这表明本次酿酒酵母培养物添加对猪肠道微生物效果非常显著。
我们观察到 ,厚壁菌门(Bacillota) 、拟杆菌门(Bacteroidota) 、螺旋体门(Spirochaetota)、假单胞菌门(Pseudomonadota)在育肥猪的肠道菌群中占有很大比例,在更低的分类水平上,每一种优势门组成都存在很大的差异。在属水平上添加组差异指示菌属是普雷沃菌属(Segatella)、厌氧弧菌属 (Anaerovibrio) 、阿加托巴克特属 (Agathobacter) 、Palleniella;差异指示种水平物种是普雷沃菌种(Segatella copri)、阿加托巴克特直肠菌(Agathobacter rectalis)和 Prevotella sp 。对照组差异指示菌属是沼泽杆菌属 (Paludibacter) 、欧文氏菌属 (Eubacterium)和 bulllifex;差异指示种水平物种是梭菌目细菌(Clostridiales bacterium)和 bulllifexporci。
添加组的显著差异的菌群中,有研究证实 Segatella copri 已被证实可参与纤维降解 ,其基因组中携带的糖苷水解酶,可分解复杂纤维,生成多种 SCFAs 。SCFAs中的丙酸又可以通过激活肠上皮 GPR41 受体,通过激活 GPR41 受体来调节肠道免疫反应和上皮细胞功能,这与添加组隐窝深度(ICD)显著降低、炎症因子 IL-6、TNF-α等显著下降结果一致。同时 Segatella copri 的 SCFAs 中的丁酸可抑制 NF-κB 通路,减少促炎因子(如 TNF-α、IL-6)释放(李林泽, 2025)。厌氧弧菌属(Anaerovibrio)是脂解作用的关键菌属,其分泌的脂肪酶可分解日粮中的甘油三酯,释放游离脂肪酸供宿主吸收(韦肖, 2022)以提升消化吸收。Agathobacter rectalis 通过丁酸激酶途径将膳食纤维发酵为丁酸,丁酸不仅是肠上皮细胞的主要能量来源,还可通过抑制HDAC(组蛋白去乙酰化酶)上调紧密连接蛋白表达,强化肠道屏障功能(Nishida, 2024)。
对照组的显著差异的菌群中,沼泽杆菌属(Paludibacter)是严格厌氧的产琥珀酸菌,其代谢产物可能通过激活 GPR91 受体促进肠道炎症(Ping, 2023),琥珀酸与 GPR91 受体结合后,启动 Gαi 和 Gαq 介导的信号通路。这些信号通路的激活会引起促炎因子的释放和免疫细胞的活化。暗示对照组肠道可能存在低度炎症状态。与对照组中 IL-6、TNF-α显著高于添加组结吻合。欧文氏菌属(Eubacterium)部分菌种(如E. hallii)可合成丁酸与上述丁酸影响途径一致。
KEGG 注释结果表明,育肥猪盲肠中最主要的微生物代谢为氨基酸代谢(Aminoacid metabolism)、碳水化合物代谢(Carbohydrate metabolism)和辅因子和维生素代谢(Metabolism of cofactors and vitamins)是丰度最高的三个通路且添加组均显著高于对照组,添加组 KEGG 一级通路上代谢(Metabolism)显著高于对照组,表明了添加组的代谢更加活跃。添加组中丰度最高的差异碳水化合物酶有 GH97、GH28、GH51、GH32、GH95、GH105和CE8。GT4、GT28、GT5,CE4 和 GH109 富集在对照组中。抗生素抗性基因丰度最高的为四环素类抗生素(Tetracycline antibiotic)其中bcrA、baeS 等在对照组中丰度较高,arlS、optrA等在添加组中丰度较高。
氨基酸代谢通路的富集主要源于 Segatella 和普雷沃菌科(Prevotellaceae)的活跃代谢。这些菌通过支链氨基酸(BCAAs)和芳香族氨基酸(AAAs)的合成(如缬氨酸、色氨酸),为宿主提供外源性氨基酸补充。
添加组 GH28 和 GH105 家族的富集,这两个家族参与着果胶与木聚糖降解能。果胶的降解产物(如半乳糖醛酸)可被 Segatella copri 等菌株进一步转化为 SCFAs 。 GH32 家族和GH97 家族,参与麸皮中菊粉和半乳糖苷类物质的代谢。GH32 通过水解菊粉β-2, 1- 糖苷键生成低聚果糖(FOS )。生成的 FOS 选择性促进双歧杆菌(Bifidobacterium)增殖,抑制大肠杆菌等致病菌(如对照组中的 Bulllifexporci),形成生态位竞争优势。这与添加组促炎因子(TNF-α、IL-6)显著降低的结果相符。对照组的GT4 家族(如脂多糖合成酶)和 GT28 家族(如肽聚糖糖基转移酶)的富集,提示对照组微生物倾向于合成细胞壁成分以应对外界压力(如基础日粮中纤维不足导致的营养竞争。CE4 家族的富集可能反映对照组微生物通过降解真菌细胞壁获取氮源。但 CE4 的过度活化可能导致宿主肠道黏膜损伤,如几丁质碎片激活 NLRP3炎症小体(刘峰, 2024),加剧肠道炎症状态。GH109 家族 (α-L-岩藻糖苷酶)的富集可能促进微生物对宿主肠道黏液层中岩藻糖基化蛋白(如黏蛋白MUC2)的降解(Bashar, 2023)。这种“黏液侵蚀 ”行为可能破坏肠道屏障,增加病原菌易位风险,解释对照组中紧密连接蛋白(ZO- 1/Occludin)表达较低的现象。对照组抗生素抗性基因 bcrA(四环素外排泵基因)和 baeS(双组分调控系统基因)的丰度较高,可能由于肠道炎症状态(如 Bulllifex 增殖)导致微生物应激响应增强。在添加组中 optrA (恶唑烷酮耐药基因)的富集,可能与 Segatella 等菌的代谢特性相关。optrA 的存在可能反映添加组微生物通过非抗生素压力(如氧化应激)激活耐药机制等,还需要进一步证明。
基于代谢组技术的组间差异代谢物比较
代谢组学是一种组学方法,用于识别和量化生物样品中存在的所有代谢物。肠道微生物群是消化的关键组成部分,在分解复杂的碳水化合物、蛋白质等营养物质的过程中会产生大量的代谢物,这些代谢物被认为是微生物区系和宿主之间的关键分子中间体,表征个体的微生物代谢特征可以全面反映微生物代谢活动的最终结果。本研究中酿酒酵母添加组中肠道代谢物与对照组出现明显分离,这与前人研究结果一致(Bai,2025)。本试验共筛选出 398 个差异代谢物,其中添加组中上调的差异代谢物有 301个,下调的有 97 个,通过对代谢物进行功能注释及富集分析,排名靠前的代谢通路有味觉转导通路(Taste transduction)、cAMP 信号通路(cAMP signaling pathway)、药物代谢-其他酶(Drug metabolism - other enzymes)、醛固酮合成与分泌(Aldosterone synthesis and secretion)、TRP 通道的炎症介质调控(Inflammatory mediator regulation of TRP channels)、脂肪酸代谢(Fatty acid metabolism)、cGMP-PKG 信号通路(cGMP-PKG signaling pathway)、核苷酸代谢(Nucleotide metabolism)、等通路均呈上调趋势。代谢物 6-deoxyerythronolide B、Adrenalone hydrochloride、Thr-Lys-Pro-Arg、 4-Pyridoxic acid 、lle-lle-Val 、Estrone 和 IVH/ICD 在多个联合分析中和显著差异菌群以及代谢通路均呈现相同趋势,暗示代谢物质之间存在协同作用。6‐脱氧红霉素 B 作为大环内酯类抗生素的前体,其上调可能提示细菌次级代谢活性增强可能会影响短链脂肪酸(SCFA)或其他代谢产物的生成,从而间接调控肠道环境。研究发现雌激素水平的变化可能会影响肠道菌群的组成,如促进特定有益菌(如乳酸菌)的生长,减少致病菌定植。调控胆汁酸代谢,进而影响脂肪的消化和吸收,可能促进短链脂肪酸(SCFA)生成,提高能量利用率。有研究表明,雌激素通过与雌激素受体(如 ERα 、 ERβ和GPR30)结合,调节肠道菌群的组成和功能。肠道菌群则通过表达β-葡萄糖醛酸酶等酶类,代谢雌激素,影响其在体内的活性和循环。这种相互作用可能对肠道健康产生影响,但具体机制仍需进一步研究。
日粮中补充该酿酒酵母培养物能够改善猪肠道微生物群落以及机体的代谢,提高厌氧弧菌属Anaerovibrio、Segatella copri、Agathobacter rectalis 等菌属和种,提高6-脱氧红霉素B(6-deoxyerythronolide B)、4-吡哆酸(4-Pyridoxic acid)、雌酮(Estrone)等有益代谢物丰度,提高猪机体的抗氧化能力,修复肠道屏障。
第五章 结论
(1)该酿酒酵母培养物可以提高育肥猪生长性能改善肠道表型。
(2)该酿酒酵母培养物可以改善育肥猪肠道免疫和屏障功能,暗示 YCB 具有改善育肥猪肠道免疫和屏障的功能。
(3)该酿酒酵母培养物可以优化育肥猪肠道微生物结构。
(4)猪肠道菌群随酿酒酵母培养物的添而发挥不同的 KEGG 新陈代谢功能。
(5)该酿酒酵母培养物调节机体代谢物,其中关键代谢物与关键差异菌、肠道表型数据极显著关联,可能是该 YCB 改善肠道代谢的生物标志物。
