

超长链鞘脂通过调节少突胶质细胞分化和膜微区完整性控制脑髓鞘形成
2026年2月,中国科学院遗传与发育生物学研究所、广州实验室、中科脂典、广州医科大学附属肿瘤医院的相关研究人员在《Journal of Translational Medicine》(2025预估IF:9.8)上发表了题为“Very long chain sphingolipids govern brain myelination by regulating oligodendrocyte differentiation and membrane microdomain integrity”的研究论文,揭示了神经酰胺合酶2(CerS2)- 超长链鞘脂轴在髓鞘发育中的关键作用,为髓鞘异常相关神经疾病的机制研究和靶向干预开辟了新方向。


CerS2合成的超长链鞘脂是脑髓鞘形成的必需分子,其缺失会使小鼠复刻脱髓鞘疾病核心表型,出现抽搐、早逝和重度髓鞘形成不足。
超长链鞘脂通过维持少突胶质细胞膜微区完整性,锚定髓鞘碱性蛋白(MBP)等关键结构蛋白,进而保障少突胶质细胞分化和髓鞘正常组装。
野生型小鼠脑内的超长链鞘脂提取物可体外挽救 CerS2 缺失导致的少突胶质细胞分化和髓鞘形成缺陷。

髓鞘是包绕在轴突外侧的富脂结构,不仅负责加快神经电信号的快速传导,还为神经元提供保护与营养支持,其损伤或形成异常会直接导致神经传导障碍,引发抽搐、瘫痪甚至死亡。多发性硬化症(MS)是最典型的脱髓鞘疾病,目前临床治疗仅能通过抗炎缓解病程,无法逆转髓鞘损伤和神经功能缺损,核心瓶颈在于髓鞘形成的分子调控机制尚未阐明。
少突胶质细胞是中枢神经系统中负责髓鞘形成的关键细胞,其前体细胞(OPCs)可分化为成熟少突胶质细胞完成髓鞘组装,是脱髓鞘疾病再生治疗的重要靶点。膜微区(脂筏)是胆固醇和鞘脂富集的细胞膜特化区域,参与细胞膜融合、蛋白定位和信号转导等关键过程,而鞘脂作为膜微区的核心组成,其代谢紊乱与脱髓鞘疾病密切相关,但具体哪种鞘脂亚型、通过何种方式调控少突胶质细胞功能和髓鞘形成,一直缺乏系统性研究。
CerS2是特异性合成 C22-C24 超长链神经酰胺的关键酶,而神经酰胺是所有复杂鞘脂的结构前体。已有临床案例显示,CerS2 基因缺失会导致患者出现进行性肌阵挛癫痫,且脑内超长链鞘脂水平显著降低,提示 CerS2 介导的超长链鞘脂代谢与神经功能密切相关。
研究团队首先构建了中枢神经系统特异性 CerS2 敲除小鼠(cKO-nestin 小鼠),结果显示该小鼠脑内 CerS2 蛋白及其合成的 C22-C24 超长链神经酰胺水平显著降低,C16 和 C18神经酰胺则出现代偿性升高,小鼠表现出体重显著低于对照组的表型,且约 4 月龄开始出现抽搐症状,胼胝体区域可见明显损伤,5 月龄后死亡率急剧升高,其中雌性小鼠的抽搐和死亡发病时间相对更早,虽未达到统计学差异但呈现出明显趋势。为明确 CerS2 缺失对小鼠神经功能的影响,研究人员开展了一系列行为学测试,发现小鼠在 Y 迷宫测试中表现出显著的空间记忆障碍,而其运动协调能力、焦虑水平、抑郁样症状及社会优势等级均无明显异常。
结合脂质组学和行为学实验结果,研究人员开展了神经病理分析,免疫荧光染色结果显示 cKO-nestin 小鼠的髓鞘碱性蛋白(MBP)表达显著降低,胼胝体厚度变薄、大脑皮质中髓鞘化轴突长度缩短,蛋白免疫印迹实验也进一步验证了小鼠胼胝体中 MBP 蛋白水平的显著下调;透射电镜的超微结构观察则发现,cKO-nestin 小鼠胼胝体区域单位面积内的髓鞘数量大幅减少,且现存髓鞘的鞘层明显变薄,证实该小鼠存在重度的髓鞘形成不足。
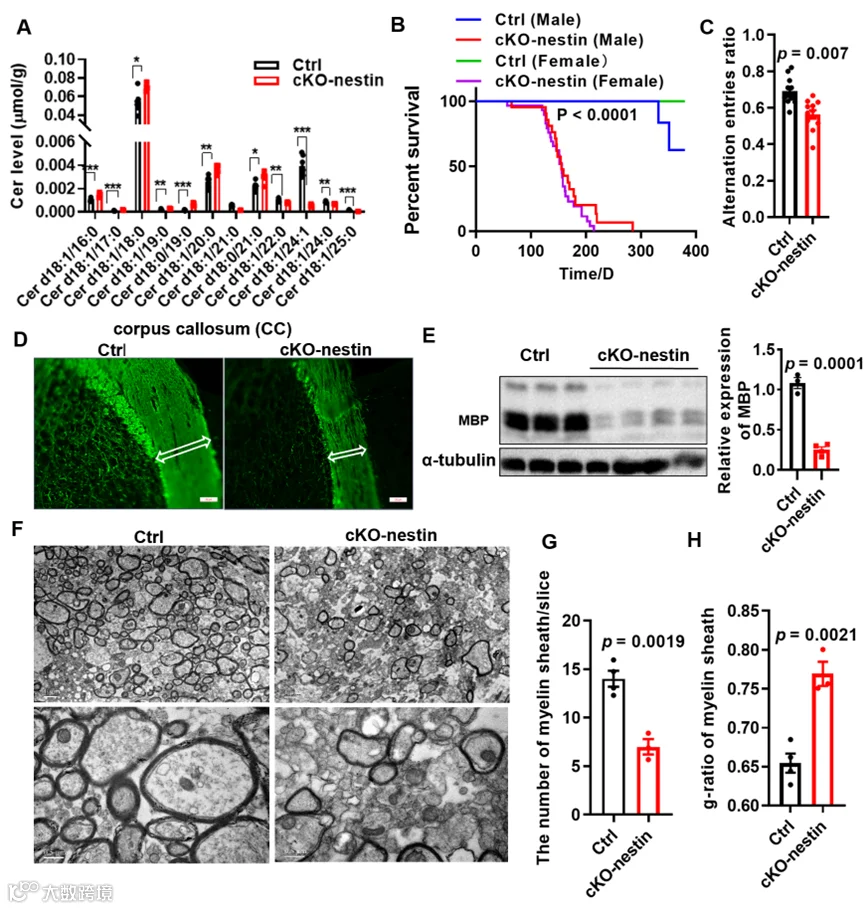
cKO-nestin小鼠的病理变化和髓鞘形成异常
考虑到小鼠髓鞘形成在出生后 15 天即开始,而 cKO-nestin 小鼠的抽搐症状在 4 月龄才出现,研究人员进一步通过透射电镜对出生后 20 天(P20)和 3 月龄的 cKO-nestin 小鼠胼胝体进行分析,结果发现早在 P20 阶段,小鼠的髓鞘数量和厚度就已显著降低,且该髓鞘缺陷在 3 月龄小鼠中持续存在并进一步加重,染色实验也证实 3 月龄 cKO-nestin 小鼠的髓鞘荧光强度和胼胝体厚度均明显下降,这表明 CerS2 敲除小鼠的髓鞘形成异常是一种早期的原发性缺陷,而非后期抽搐引发的继发性结果。
为确定髓鞘异常的核心细胞来源,研究人员构建了少突胶质细胞特异性 CerS2 敲除小鼠(cKO-OL),该小鼠完全复刻了 cKO-nestin 小鼠的核心表型,包括体重降低、抽搐、死亡率升高以及髓鞘数量减少、鞘层变薄的髓鞘形成障碍,这一结果证实少突胶质细胞中的 CerS2 缺失是驱动髓鞘病理改变的关键因素。
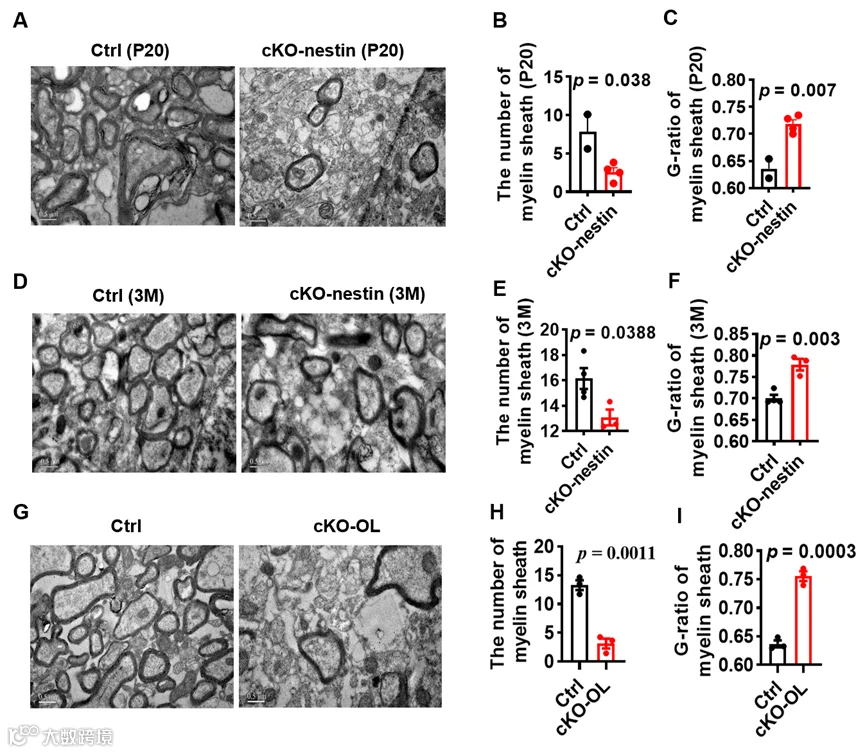
cKO-nestin和cKO-OL小鼠髓鞘形成异常的电镜检查
鉴于 CerS2 是超长链神经酰胺的主要合成酶,研究人员对 cKO-nestin 小鼠的脑内脂质组进行了全面分析,结果显示除超长链神经酰胺外,其下游对应的超长链鞘磷脂(SM)以及硫苷脂(SL)、半乳糖神经酰胺(GalCer)、葡萄糖神经酰胺(GluCer)等复杂糖鞘脂水平均显著降低,而短链鞘脂则呈代偿性升高,同时小鼠脑内的鞘氨醇 - 1 - 磷酸(S1P)各亚型水平也显著下调。因此,cKO-nestin 小鼠脑内的大规模脂质重构主要由极长链神经酰胺的减少引发,极长链神经酰胺是维持脑内脂质整体结构的关键枢纽,而该结构对髓鞘形成至关重要。
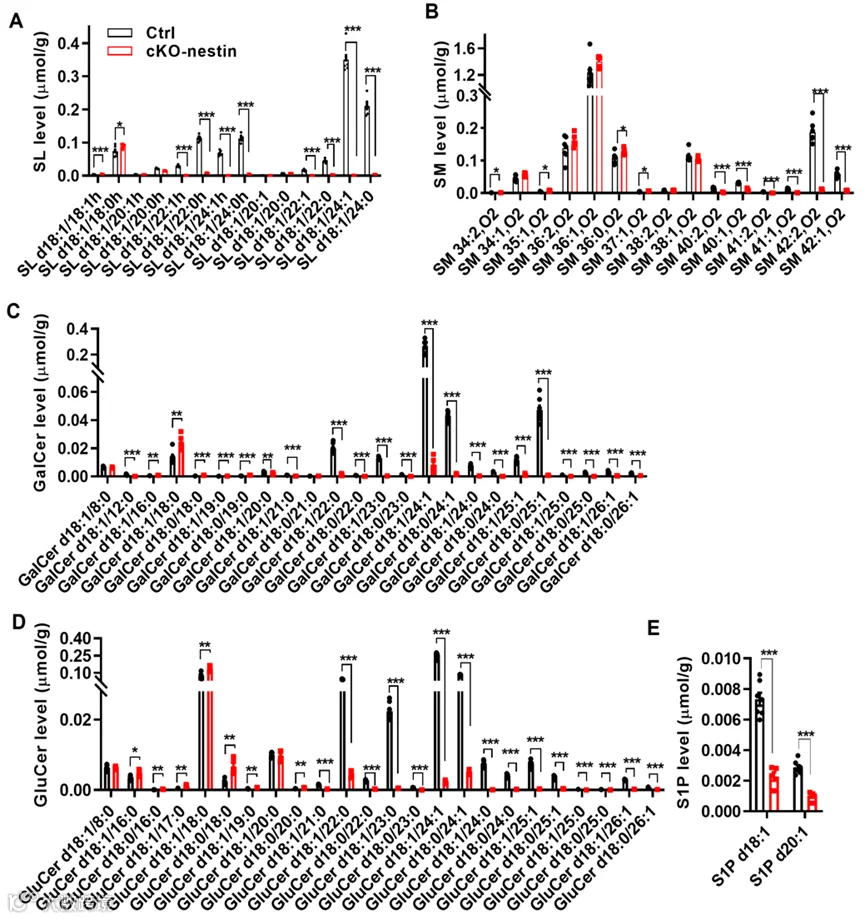
cKO-nestin小鼠的脂质改变
为进一步明确 CerS2 缺失引发的脂质变化在脑内的空间分布特征,研究人员采用空间分辨率达 10 μm 的解吸电喷雾电离质谱成像(DESI-MSI)技术对小鼠脑片进行分析,发现 SM 42:1;O2、己糖神经酰胺(HexCer)40:1;O2 等关键超长链鞘脂的缺失并非全身性,而是特异性定位于胼胝体、内外囊等脑内主要白质纤维束区域,且在海马伞、髓纹等其他髓鞘化区域,超长链鞘脂的荧光强度也显著下降,该空间分布特征在基质辅助激光解吸电离质谱成像(MALDI-MSI)的独立实验中得到了重复验证;与之相反,Cer 36:1;O2、SM 34:1;O2 等短链鞘脂的升高则主要出现在纹状体、苍白球和海马区域,二者的空间分布差异直接证实了超长链鞘脂缺失与髓鞘损伤区域的精准共定位。
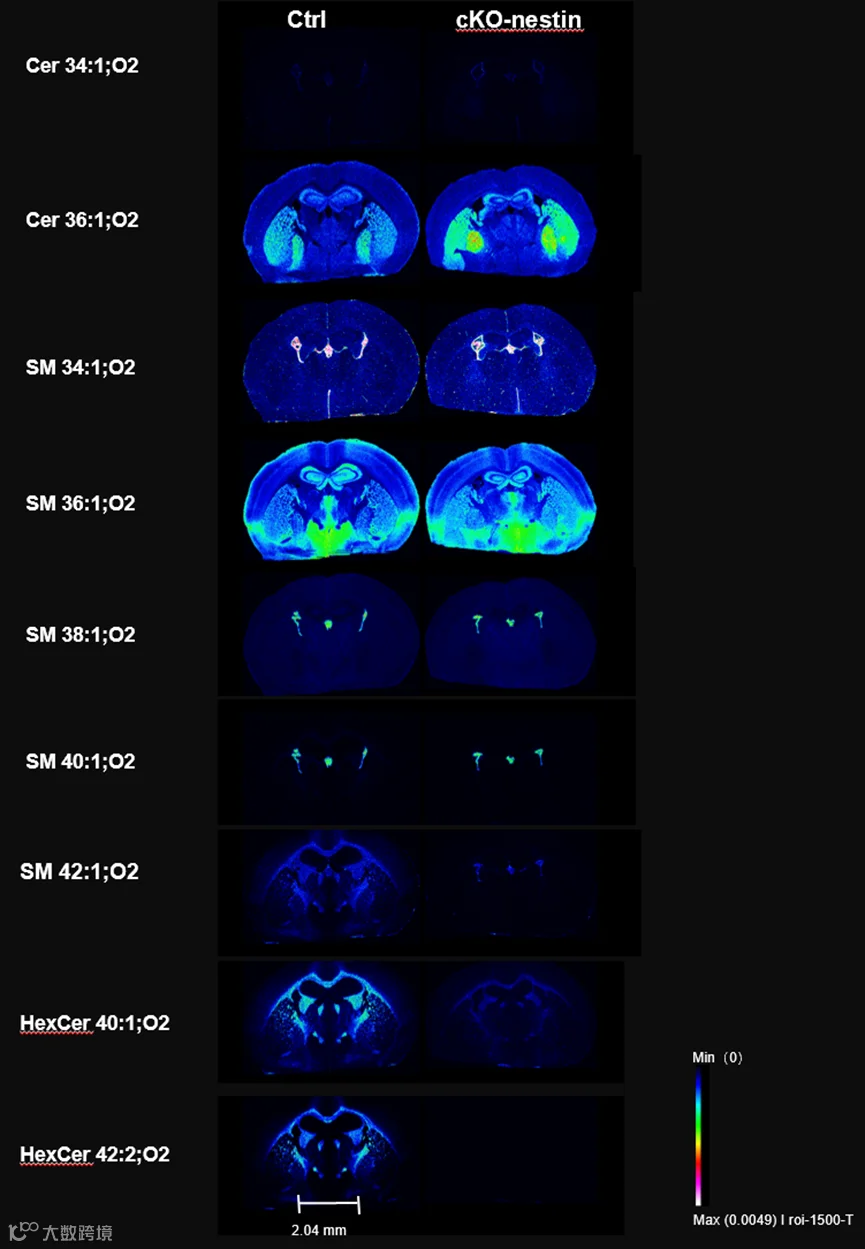
Ctrl和cKO-nestin小鼠脑区的空间脂质分布
为解析 CerS2 - 超长链鞘脂轴调控髓鞘形成的分子机制,研究人员对 3 月龄雄性 cKO-nestin 小鼠的脑内蛋白组进行了分析,基因本体(GO)富集结果显示,小鼠脑内与细胞膜融合、细胞器膜融合相关的通路显著下调,而这两个通路是中枢神经系统髓鞘鞘层组装的核心分子事件。进一步分析脂筏组分的蛋白质组发现,髓鞘核心结构蛋白 MBP 在脂筏中的含量显著减少,同时蛋白脂质蛋白 1(PLP1)、髓鞘相关糖蛋白(MAG)也呈共降低趋势,而髓鞘少突胶质细胞糖蛋白(MOG)水平无明显变化,该蛋白变化模式与髓鞘形成不足的表型高度契合,免疫印迹实验进一步验证了 cKO-nestin 小鼠脂筏组分中 MBP 蛋白水平的显著降低,提示超长链鞘脂是维持膜微区完整性的关键,其缺失会导致脂筏结构破坏,进而引发髓鞘关键蛋白的定位异常。
此外,研究人员还构建了背根神经节(DRG)神经元 - 少突胶质细胞前体细胞共培养的体外髓鞘形成模型,进一步验证膜微区的功能,结果发现 CerS2 敲除的少突胶质细胞形成的髓鞘节段显著短于对照组;而向野生型共培养体系中加入 β- 环糊精(β-CD,特异性破坏膜微区的试剂)后,野生型少突胶质细胞的髓鞘形成也被显著抑制,且 50 μM 和 250 μM 浓度下的抑制效果无显著差异,证实膜微区完整性是髓鞘形成的必要条件,其破坏可直接引发髓鞘形成障碍,与 CerS2 缺失导致的表型一致。
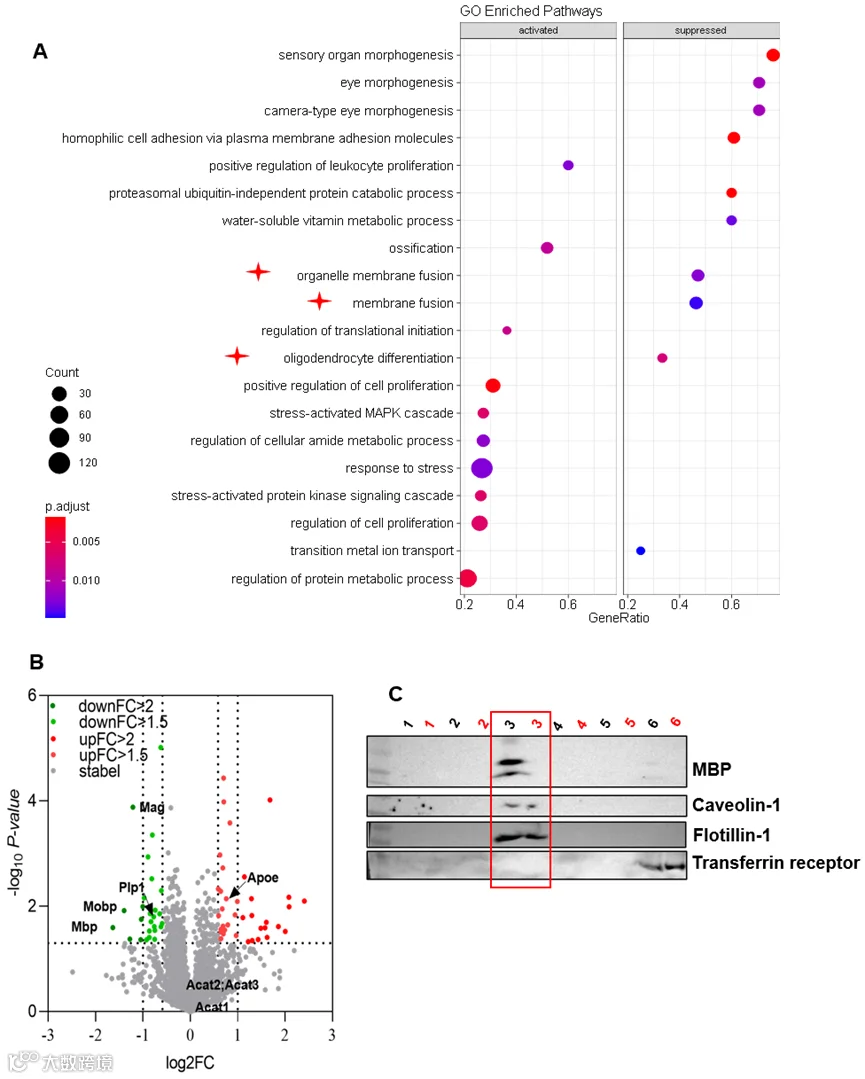
cKO小鼠的微结构域改变
脑蛋白质组学分析还提示 cKO-nestin 小鼠脑内少突胶质细胞分化相关通路下调,研究人员通过体外分化实验验证发现,CerS2 敲除的神经前体细胞分化为成熟少突胶质细胞(MBP + 细胞)的比例显著低于对照组,证实超长链鞘脂参与调控少突胶质细胞的分化。为进一步明确超长链鞘脂的功能,研究人员开展了挽救实验,向敲除的神经前体细胞中补充野生型小鼠脑脂质提取物(含正常水平超长链鞘脂),可显著恢复其分化能力,且在 DRG 神经元共培养体系中,其髓鞘形成能力也得到明显提升;而补充 cKO-nestin小鼠的脑脂质提取物(缺乏超长链鞘脂)则无此效果。
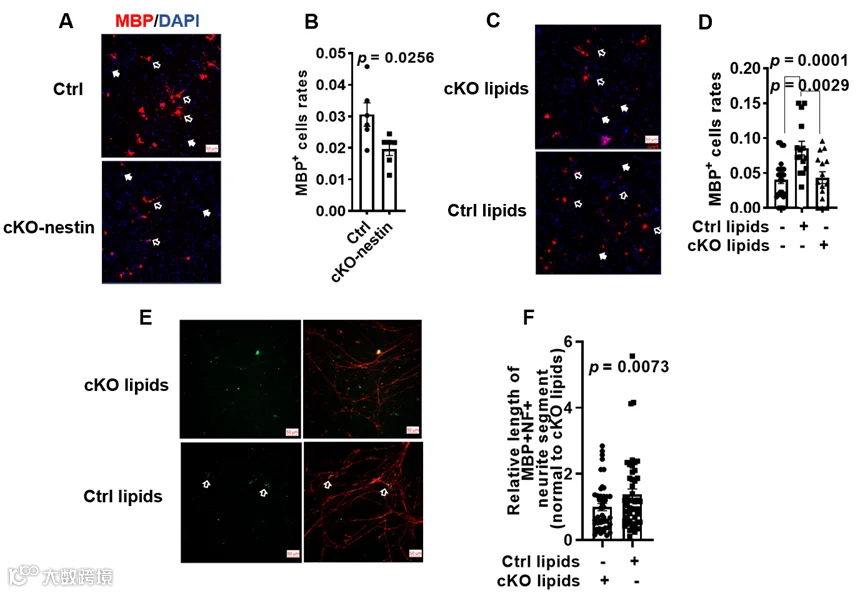
超长链鞘脂对OL分化和髓鞘形成的影响
综上所述,该研究结果不仅确立了 CerS2 缺陷与发育性髓鞘形成失败之间的因果关系,还为未来探究极长链鞘脂补充作为相关神经系统疾病治疗策略的研究奠定了重要基础。

Science Translational Medicine:神经发育中的脂质重塑,揭示神经发育障碍疾病相关分子机制
Cell Metabolism:糖鞘脂:连接葡萄糖代谢与抗癌免疫的关键,让CD8⁺ T细胞杀瘤更高效
JGG:税光厚团队长文综述鞘脂在病理生理过程中的新作用及治疗潜力
Clin Transl Med:税光厚团队揭示CerS2介导Mad2信号通路维持肝细胞正常分裂的分子机制

点击“阅读原文”即可查看或下载原文。