
作物对磷的吸收、运输和利用
作物对磷的吸收、运输和利用

磷是植物生长发育不可缺少的营养元素之一,它既是植物体内许多重要有机化合物的组分,同时又以多种方式参与植物体内各种代谢过程。磷对作物高产及保持品种的优良特性有重要作用。

吸收
土壤溶液中的磷浓度为0.5~2umol/L,而木质部汁液含磷量为0.4mmol/L,是土壤中磷的浓变的400倍。同样,根细胞中磷酸盐的浓度显著高于土壤溶液中的磷,说明植物根系能逆浓度梯度主动吸收磷酸盐。植物吸收磷的主要形态是不同的磷酸根。
植物根系主要在根毛区吸收磷,这是因为大量根毛提高磷的吸收面积,且根毛区的木质部发育成熟,有利于磷向地上部运输。而根尖分生区、伸长区因木质部未发育完全,不利于磷的吸收和运输。
磷的主动吸收过程发生在根表皮细胞质膜上。质膜上的H+-ATPase水解ATP形成质子驱动力,H2PO-4与磷转运蛋白结合后进入细胞膜内。植物体内存在高亲和、低亲和两种磷吸收系统。缺磷时,质膜上高亲和的磷酸盐转运蛋白转运磷(Km值在mol/L范围);磷充足时,低亲和的磷酸盐转运蛋白转运磷(Km值在mmol/L范围)。负责根系吸收磷的是PHT1磷转运蛋白家族。这类磷转运蛋白一般由12个跨膜结构域组成,属于H2PO-4/H+共转运蛋白(图5-10)。PHT1家族的磷转运蛋白大多数在根系中表达,受缺磷诱导。目前,在小麦、水稻、玉米、番茄、土豆、大豆等植物中,已鉴定出来编码PHT1家族的磷转运蛋白基因。此外,在叶绿体和线粒体等细胞器的被膜上,同样鉴定到磷转运蛋白家族成员,介导磷从细胞质向这些细胞器中的转运。依据功能和亚细胞定位特征,植物中的磷转运蛋白大体划分为 PHT2,PHT3,PHT4 和 PHT5家族。
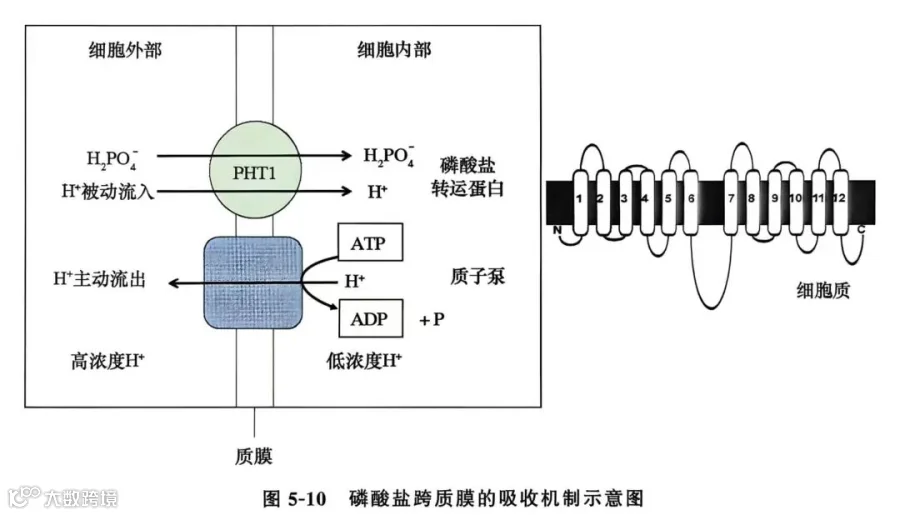
根系吸收的磷酸盐进入细胞后迅速参与代谢过程,并通过共质体途径运输到内皮层后装载进入木质部,然后磷向地上部运输,供给植株生长。研究表明,磷酸盐转运蛋白PHO1负责无机磷向木质部装载,其主要在中柱鞘和木质部薄壁细胞中表达(Hamburger et al.,2002)。在木质部导管中运输的磷绝大部分是无机态磷酸盐,有机磷含量极少;韧皮部中的磷包括无机磷和有机磷。
植物体内磷的移动性较大,可以通过韧皮部双向运输,在某些情况下,向地上部运输的磷中,约有50%以上的磷可通过韧皮部再转移至植物体内其他部位,尤其是快速生长的器官。大麦根基部吸收磷酸盐后,其被转移到根尖和植物顶部。同时,新叶中的磷一部分来自根系吸收的磷酸盐,另一部分来自老叶中有机磷释放出的无机态磷。在成熟期,禾谷类作物中60%~85%的磷素转移至籽粒。大豆籽粒开始形成后,由根部吸收供应的磷大约占总磷量的45%,其余磷来源于营养器官的再利用。
尽管植物细胞中的含磷化合物存在着分隔现象,但植物各部位和不同形态的磷可以相互转化和调节。磷进人细胞以后,一部分用于合成磷脂、DNA和RNA;一部分用于合成能量物质ATP;一部分以无机磷形态存在于液泡中。植物细胞中的无机磷主要储存于液泡当中,液泡中磷素的储存和输出对于维持植物中的磷素平衡和植物的生长发育非常重要。当外界磷素充足时,植物吸收的大部分无机磷会储存在液泡中;当外界磷素缺乏时,液泡储存的无机磷会被释放到胞质中供植物利用。目前已经发现SPX一MFS家族的磷转运蛋白可能参与了液泡无机磷的运输。
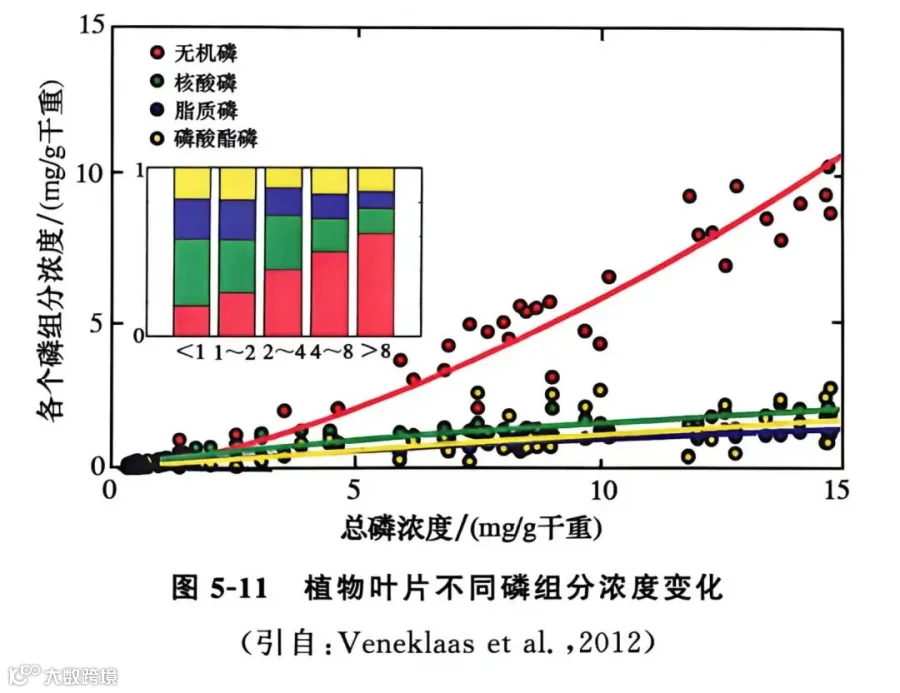
成熟叶片所需要的养分,主要靠蒸腾作用将根系吸收的养分经木质部运输到地上部,再通过茎节将养分运输分配到不同的叶片中。由于未展开的新生叶片的蒸腾作用较弱,因此,新叶所需要的养分主要源于老叶中养分的再利用。植物叶片中的磷库大致可分为4个主要组分,即无机磷(Pi)、磷酸酯类磷(phosphorus esters)、脂磷(phosphorus lipids)核酸磷(nucleic acids)。各磷库的大小依次为无机磷>核酸磷>脂质磷>磷酸酯类磷(图5-11)。在叶片中,核糖体RNA磷占总磷含量的约30%,核糖核酸酶(RNase)水解衰老叶片中的核糖体RNA,释放出的磷运输到新叶,供给植物生长。与新叶相比,在老叶中磷脂的含量较低,但双半乳糖甘油二酯、硫代异鼠李糖甘油二酯含量较高,释放出的磷供给新叶的生长。
土壤中的磷易被固定、磷扩散系数低(为10-10~10-cm2/s)。为了获取土壤中的磷,植物进化出了一套精密的信号感知系统。首先,根系感受细胞外磷水平的变化,然后通过木质部将信号传递给叶片,叶片感受到根传递的信号后,通过韧皮部再将信号传递到茎的顶端及根系中,调节根系对磷的吸收,以及植株生长发育。
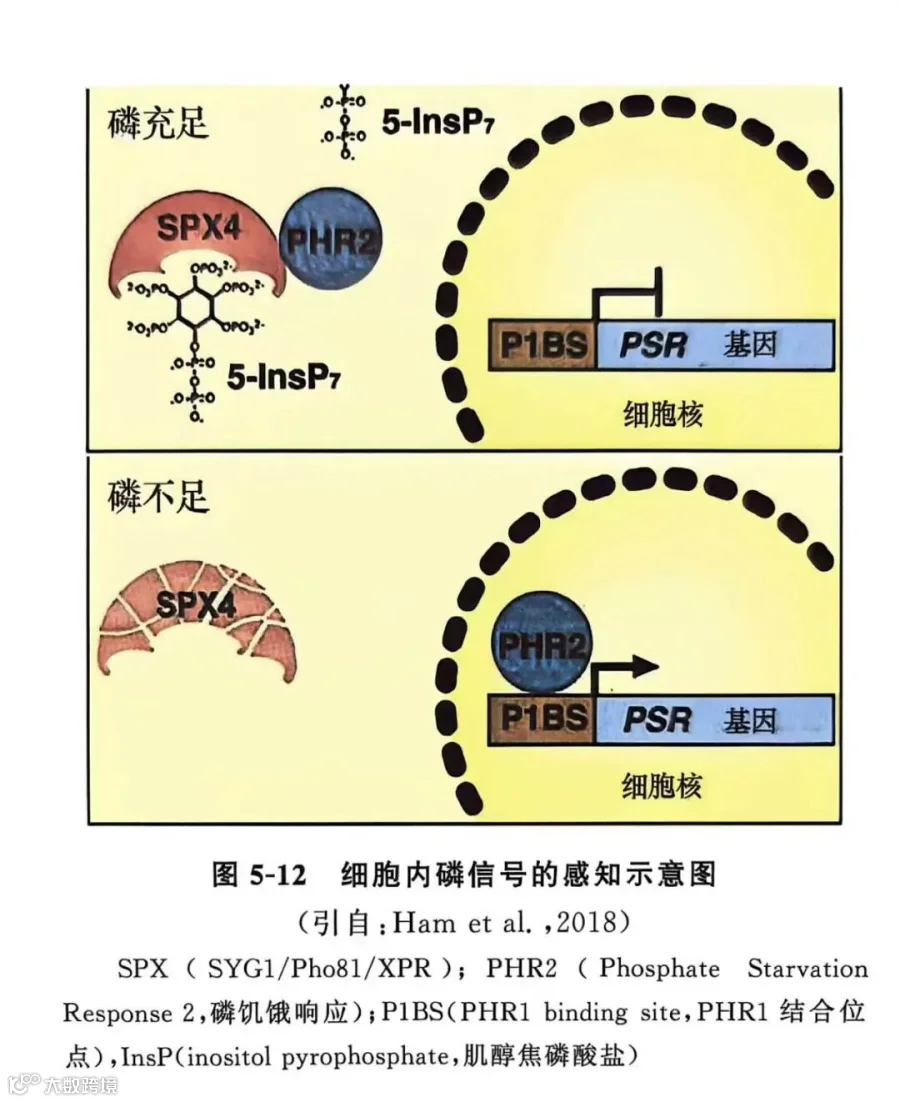
植物根尖是感受缺磷的部位。目前认为,植物通过两条途径感受介质中的磷水平,一是位于根细胞膜的感受器,其感受外部磷水平;二是细胞内的传感器,感受内部磷水平。当植物根尖处于低磷环境时,位于质膜上的感受器就会产生或激发信号,然后根尖细胞内的感受器产生胞内缺磷信号。在水稻和拟南芥中,含SPX结构域的蛋白可能是植物体内的磷感受器(Puga et al.,2014;Wang et al.,2014)。磷饥饿响应PHR在植物磷信号调控网络中作为中心调控因子,通过结合顺式作用元件P1BS调控植物磷信号。肌醇焦磷酸盐(InsP)的含量随细胞内磷水平的变化而变化。如图5-12所示,供磷充足时,InsP直接结合磷的受体SPX,促进SPX和PHR的相互作用,以抑制PHR对缺磷响应基因的激活;缺磷时,InsP含量降低,SPX4不能和PHR结合,PHR结合到P1BS位点,激活缺磷响应基因的表达,启动植物对低磷胁迫的应答机制。
植物体内的缺磷信号物质主要包括激素、糖miRNA等。植物根尖感知磷信号并传递到叶片细胞分裂素和独脚金内酯等在根系向叶片的磷信号网络中起着重要作用。研究表明,细胞分裂素抑制植物对缺磷的响应。例如,外源施用细胞分裂素后显著抑制了缺磷响应基因的表达。独脚金内酯主要在根系中合成,抑制腋芽的形成和发育,从而控制其分支。拟南芥缺磷时,根内合成独脚金内酯的数量显著上升,在木质部汁液中检测到独脚金内酯,最终调控植株地上部的分枝。韧皮部转运的RNAs、蛋白质、糖和其他代谢产物在地上部到地下部传递的信号中也起着重要作用。研究发现,由miR399和miR827介导磷信号从植物地上到地下的传递。
植物吸收磷受很多因素的影响。其中有植物生物学特性和环境条件两个方面,尤其是植物的吸收能力。
1.作物特性
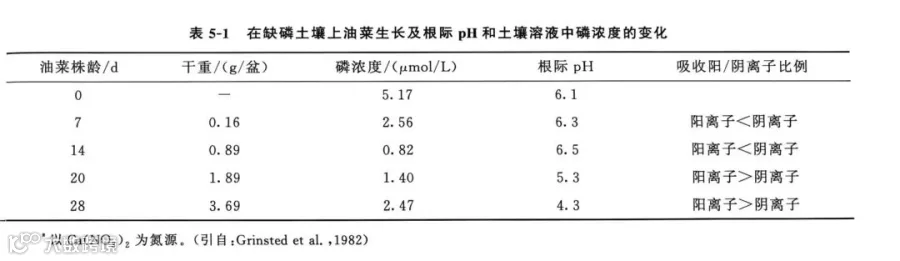
不同植物种类,甚至不同的栽培品种,对磷的吸收都有明显的影响。例如,植物根系形态不同,根系改变局域土壤pH的能力也不相同。不同植物的根系,其密度、形状、结构等特性都有差异,因此吸收能力明显不同,尤其是土壤溶液中磷浓度很低时,更是如此。根毛对植物吸收磷有明显作用,洋葱因为没有根毛,其吸磷能力就比较弱。油菜的根系并不发达,也不能感染菌根,但它吸磷的能力却较强。究其原因,是油菜在缺磷的情况下,根系能自动调节其阴阳离子吸收的比例,使根际土壤酸化,从而提高土壤溶液中磷的浓度(表5-1)。
许多植物的根系具有分泌H和有机酸的能力。最典型的例子是白羽扇豆,它除了能分泌H”外,还能分泌大量的有机酸,如柠檬酸,能螯合铁、铝离子,从而提高了根际土壤磷的有效性。白羽扇豆在缺磷时,还能形成排根,通过扩大根系表面积以增加对磷的吸收。
2.土壤供磷状况
植物主要是利用土壤中的无机态磷。虽然植物可吸收少量有机态磷,然而,有机磷必须转化为无机态磷后才能被植物吸收。土壤溶液中磷酸根离子的浓度很低,其移动方式主要靠扩散作用。扩散作用则与土壤固相-液相以及液相-液相之间的平衡有关。影响磷酸根离子扩散的因素很多,除磷酸根离子浓度外,其他因素如土壤的温度、水分、质地、孔隙度和黏粒矿物种类等均影响磷的扩散系数。通常温度升高、水分增加、土壤松散均有利于无机磷的扩散作用。
3.菌根
菌根能增加植物吸磷的能力。菌根根外庞大的菌丝扩大了根系的吸收面积,菌丝对磷酸盐亲和力较高,此外菌根菌丝分泌质子和有机酸阴离子也能促进难溶性磷的溶解,磷酸酶则能矿化土壤中的有机磷。不同植物对菌根依赖性不同,例如,小麦对菌根真菌依赖性较低,玉米为菌根依赖性较高的作物。除宿主植物外,菌根对植物磷吸收的贡献与土壤磷的供应强度密切相关,低磷时菌根促生,反之高磷时抑制植物生长。
4.环境因素
环境条件中以温度和水分的影响最为明显。土壤温度是影响根系吸收磷的重要因素。在一定范围内(10~40°C),提高土温可增加植物对磷的吸收。土温升高后,不仅土壤溶液中的磷扩散速度加快,而且根和根毛生长速度也相对加快,根的呼吸作用明显加强,这些均有利于促进植物对磷的吸收。增加水分促进了磷在土壤中的扩散,提高磷的生物有效性。
5.养分的相互关系
磷与氮在植物吸收和利用方面有相互影响。施用氮肥常能促进植物对磷的吸收利用。因为磷参与氮代谢、硝酸盐还原、氨的同化以及蛋白质合成。氮磷配合施用可促进植物生长得更好,反过来又促进植物吸收更多的氮和磷。
遵循作物吸磷规律科学施磷
活化土壤有效磷,提升养分转化率
夯实作物高产基础



青岛远东联盟进出口贸易
有限公司
文字:小戴
编辑:悦悦
图片:小苏
复审:Connie Lin
终审:Marcel Wang
来源:Plant Nutrition
声明:以上内容仅供学习交流使用,不具有任何商业用途
版权归原作者所有,如有问题请及时联系我们以作处理
本内容不做商用
部分图片来源于网络
如有侵权,请联系删除