

Section.01
mTOR:生命调控的 "总指挥"
mTOR(哺乳动物雷帕霉素靶蛋白)是细胞内最核心的信号枢纽之一,它像一位精明的"代谢总开关",时刻监测着环境中的营养、能量和生长因子信号,决定细胞是该"疯狂生长"还是"节能自保"[1]。简单来说:
营养充足时:mTOR 开启,促进蛋白质、脂质、核苷酸合成,驱动细胞增殖
营养匮乏时:mTOR 关闭,启动自噬(细胞自我清理),回收资源维持生存
Section.02
核心拆解:
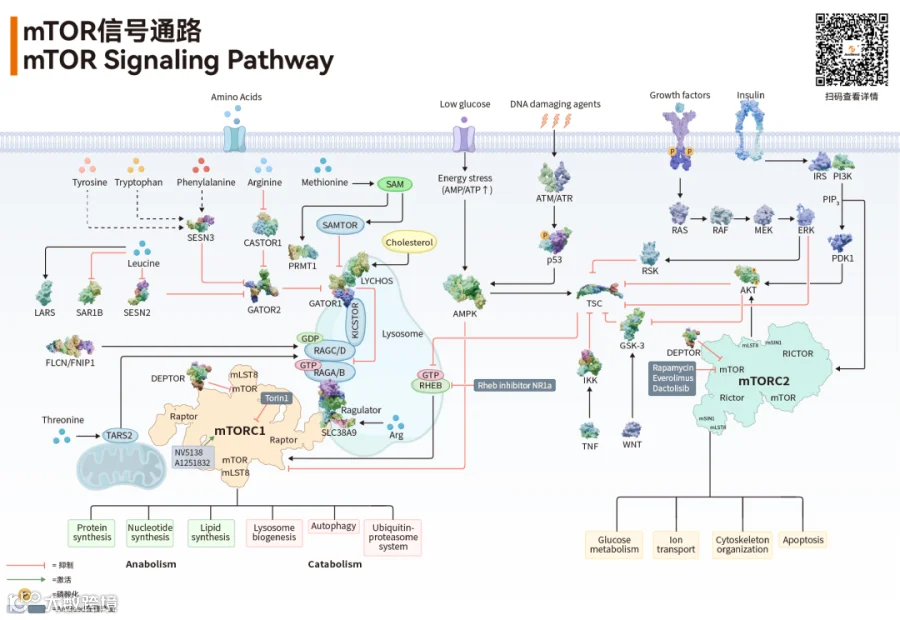
mTOR 的 “双引擎”——mTORC1 vs mTORC2
mTOR 并非单一蛋白,而是作为催化核心,分别与 Raptor 或 Rictor 等适配蛋白组装成功能特异的mTORC1 与 mTORC2 复合体,二者在底物识别、信号响应及下游效应上分化明显、协同整合,共同驱动细胞代谢的"双引擎"[2,3]

近年研究证实,mTORC1 在溶酶体膜通过 IRTKS 介导的液 - 液相分离(LLPS)形成动态凝聚体,呈脉冲式节律激活;其组装紊乱致自噬受阻、衰老加速,为间歇性调控优于持续抑制提供分子依据[4]。
Section.03
上游调控:
四大信号,精准控制 mTOR 开关
mTOR 并非 “随心所欲”,而是受四大上游信号层层调控,形成精密的 “传感 - 调控” 网络。
1.营养信号:氨基酸、葡萄糖的 “传感机制”
氨基酸通过 SESTRIN2-GATOR2 和 CASTOR1-GATOR1 等传感器网络激活 RAG GTP 酶,将 mTORC1 招募至溶酶体膜;葡萄糖经 AMPK 间接调控,脂肪酸则通过 FABP5 直接激活。2024 年《Nature Communications》揭示,果蝇与脊椎动物独立演化出结构迥异的 SAM 传感器,却通过 GATOR2 枢纽实现功能趋同,体现 mTORC1 通路整合新营养传感模块的进化可塑性[5]。
2.能量信号:AMPK——mTOR 的 “天然刹车”
AMPK 作为细胞能量状态的核心传感器,在 ATP 不足、AMP/ADP 升高时被激活,直接磷酸化 TSC2 和 Raptor,强力抑制 mTORC1 活性,从而启动自噬、抑制合成代谢,实现"节约能量、保命优先"的应激响应。这一机制是连接能量代谢与细胞生长调控的关键节点,也是二甲双胍等药物间接抑制 mTOR 的分子基础[1]。
3.生长因子:胰岛素 / IGF-1 的 “激活通路”
胰岛素/IGF-1 结合受体后激活 PI3K,生成 PIP3 招募并激活 AKT;AKT 通过磷酸化 TSC2 解除其对 RHEB 的抑制,从而激活 mTORC1,同时 mTORC2 磷酸化 AKT 的 Ser473 位点形成正反馈,构成"生长-代谢"联动网络。这一轴心是 PI3K/AKT/mTOR 通路的核心,也是癌症中该通路过度激活的主要驱动机制[1]。
4.压力信号:DNA 损伤、氧化应激的 “抑制开关”
细胞遭遇 DNA 损伤、氧化应激或缺氧时,优先抑制 mTOR 以启动修复程序:DNA 损伤通过 ATM/ATR 激酶磷酸化 TSC2 和 p53 抑制 mTOR;氧化应激通过 ROS 升高激活 AMPK 间接抑制 mTOR;缺氧则通过 HIF-1α 诱导 DDIT4(REDD1)表达,解除 RHEB 对 mTORC1 的激活。这种"生长让位于修复"的优先级切换,是细胞在应激条件下维持基因组完整性和存活的关键策略[1]。
Section.04
下游功能:
mTOR,掌控细胞生死代谢的 “全能王”
mTOR 激活后,通过三大核心下游功能,全面调控细胞命运,堪称细胞的 “全能调控师”[1]。
(一)代谢调控:细胞的 “营养分配官”
蛋白质合成:磷酸化 S6K1 与 4E-BP1,启动核糖体翻译,是肌肉生长与组织修复的核心。
脂质/核苷酸合成:激活 SREBP 与 CAD 酶,为细胞增殖提供膜结构与 DNA 原料。
自噬抑制:营养充足时关闭自噬。长期不“关机”,细胞垃圾堆积,衰老加速。
(二)线粒体与肠菌互作:跨系统调控的“能量-微生态”枢纽
线粒体不仅是细胞的“能量工厂”,更是 mTOR 感知代谢状态、协调全身稳态的核心枢纽。近年研究揭示,mTOR 对线粒体的调控已从单一的“产能管理”升级为与肠道微生态深度联动的“跨系统网络”。
1. 线粒体生成
mTORC1 通过 PGC-1α、YY1 驱动线粒体生物合成
增加数量与嵴密度,匹配快速生长的能量需求
2. 质量控制与氧化应激
促进线粒体融合、抑制过度分裂,维持功能完整
营养充足时抑制 PINK1/PARKIN、BNIP3 介导的线粒体自噬
生理性低 ROS 促生长;mTORC1 持续亢进则过度产生 ROS,削弱 NRF2 抗氧化通路
3. 肠上皮-菌群双向轴[6]
线粒体稳态并非孤立存在,而是与肠道微生态形成闭环反馈:
肠道上皮 mTORC1 调控黏液层厚度与抗菌肽(如 Reg3γ)分泌
共生菌代谢物(丁酸、吲哚衍生物)通过 HDAC 抑制/AMPK 激活 负反馈抑制 mTORC1
高脂/高糖饮食破坏该环路,导致肠屏障渗漏与全身代谢性炎症
(三)免疫调控:免疫细胞的 “代谢编程师”
mTOR 决定免疫细胞分化、功能、命运,是免疫代谢的核心。
1. T/B 细胞分化:决定免疫应答方向
mTORC1:促进Th1、Th17、效应 CD8+T 细胞分化,增强炎症;
mTORC2:促进Th2、Tfh 细胞分化,调控体液免疫;
抑制 mTOR:促进 调节性 T 细胞(Treg) 分化,诱导免疫耐受。
2. 巨噬细胞极化:炎症 vs 修复
mTORC1:促进M1 巨噬细胞极化,释放炎症因子;
mTORC2:促进M2 巨噬细胞极化,介导修复、抗炎。
3. 免疫衰老:老年免疫力衰退
mTOR 长期激活:免疫细胞代谢紊乱、增殖能力下降、记忆丧失;
抑制 mTOR:恢复免疫细胞功能、增强疫苗应答、延缓免疫衰老。
Section.05
疾病关联:
mTOR 失衡,百病丛生
癌症:PI3K/Akt/mTOR 是实体瘤高频异常通路之一。PTEN 缺失、Rheb 或上游受体突变可使 mTORC1 持续激活,驱动增殖、血管生成与免疫逃逸。单药易触发代偿通路,当前临床前沿聚焦联合策略(如联用免疫检查点抑制剂、靶向药或自噬调节剂)[7]。
衰老 :美国 NIA 干预测试计划(ITP)及多项独立动物研究表明:间歇性、低剂量、短周期的 mTOR 调节在延长健康寿命的同时,可显著降低持续给药带来的代谢与免疫副作用。机制可能涉及周期性自噬激活、衰老相关分泌表型(SASP)下调及组织特异性稳态维持。目前“组织靶向”与“给药节律”已成为转化研究重点[8]。
代谢:mTORC1 过度激活可通过 S6K1-IRS1 负反馈环路诱发胰岛素抵抗。肝靶向或纳米递送 mTORC1 调节策略在临床前脂肪肝/NASH 模型中显示出同步改善脂质代谢与炎症的潜力[9]。
神经疾病:阿尔茨海默病等蛋白病中,mTOR 亢进与自噬流受阻、异常蛋白累积相关;结节性硬化症(TSC)与部分难治性癫痫属于典型 mTOR 通路疾病。脑穿透型低剂量调节剂的探索正在进行中[10]。
Section.06
靶向治疗:
从实验室到临床,mTOR 药物全解析
第一代:mTORC1 变构抑制剂
雷帕霉素及其衍生物(如依维莫司、替西罗莫司)通过变构方式部分抑制 mTORC1,却无法彻底阻断 4E-BP1 磷酸化,且会解除 mTORC1 对 mTORC2 的负反馈抑制,导致 PI3K/AKT 信号通路 代偿性激活,促进细胞存活与耐药;目前均已上市,用于肾移植抗排斥、肾癌、乳腺癌及神经内分泌瘤等[1]。
第二代:mTORC1/2 双重激酶抑制剂
直接靶向 mTOR 激酶域同时抑制两个复合物,阻断 AKT 反馈激活,但毒性叠加导致治疗窗口窄;Sapanisertib 处于 II 期(子宫内膜癌、肾癌、乳腺癌),Onatasertib 处于 I/II 期(晚期实体瘤)[11]。
第三代:mTORC1 选择性抑制剂
精准区分 mTORC1 与 mTORC2,只关"生长引擎"不动"存活引擎",力图保留疗效的同时剥离代谢副作用,为结节性硬化症(TSC)等 mTOR 病的长期管理提供"减毒增效"方案。目前大多处于早期临床阶段[12]。
第四代:多节点抑制剂
不再纠结于单一靶点的抑制深度,转而通过同时打击 PI3K 和 mTOR 或多个通路节点,从源头掐断反馈激活和旁路耐药,通过低剂量联合用药,达到比高剂量单药更强、更彻底的抑制效果[13]。代表药物 Gedatolisib 作为 PI3K 和 mTOR 的双重可逆抑制剂,基于 ER+/HER2- 乳腺癌(CDK4/6 治疗后进展)和子宫内膜癌的 I/II 期数据[14],Sapanisertib 联合 PI3Kα 抑制剂 Serabelisib 的方案目前处于 I/II 期临床[15]。
📚 参考文献
1.Signal Transduct Target Ther. 2025;10(1):428.
2.Nature. 2025;647(8089):536-543.
3.Science. 2026;391(6789):eadv7111.
4.Cell Rep. 2026;45(2):116893.
5.Nat Commun. 2024;15(1):2517.
6.Front Gastroenterol (Lausanne). 2024;3:1386069.
7.Cancer Res. 2022;82(21):3884-3887.
8.Mol Metab. 2024;81:101902.
9.Nat Rev Mol Cell Biol. 2023;24(12):857-875.
10.Nat Commun. 2022;13(1):4665.
11.Signal Transduct Target Ther. 2025;10(1):198.
12.Frontiers in neuroscience vol. 19 1595880.
13.British journal of cancer vol. 133,2 (2025): 144-154.
14.Int. J. Mol. Sci. 2023, 24, 4522.
15.Gynecologic oncology vol. 166,3 (2022): 403-409.
| AmBeed产品推荐 |
|
|
|
|






往期推荐
不止于经典:解密cAMP信号,洞悉其在癌症中的多重调控与治疗新靶点
宫颈癌的“幕后黑手”:HPV是如何一步步“策反”我们身体的?