
摘 要
胰岛素抵抗是代谢综合征和2型糖尿病的主要病理生理学基础 。以前的宏基因组学研究已经描述了肠道菌群的特征及其在胰岛素抵抗中代谢主要营养物质的作用-。特别是,共生菌的碳水化合物代谢被认为是可以为宿主提供高达10%的总能量,从而在肥胖和糖尿病前期的发病机制中起作用。然而,其潜在的机制仍不清楚。在这里,我们使用一种全面的多组学策略来研究人类中这种关系。我们结合无偏倚的粪便代谢组学和宏基因组学、宿主代谢组学和转录组学数据,来描绘肠道菌群在胰岛素抵抗中的参与情况。这些数据显示,粪便中的碳水化合物,特别是宿主可利用的单糖,在胰岛素抵抗的个体中增加,并与微生物的碳水化合物代谢和宿主的炎症细胞因子相关。我们鉴定了与胰岛素抵抗和胰岛素敏感性相关的肠道细菌,并发现它们显示出不同的碳水化合物代谢模式,并证明了胰岛素敏感性相关的细菌可以改善小鼠模型中的胰岛素抵抗的宿主表型。我们的研究,为胰岛素抵抗中的宿主-微生物关系提供了一个全面的视角,揭示了微生物的碳水化合物代谢的影响,为改善胰岛素抵抗提供了一个潜在的治疗靶点。
正 文
我们分析了306名(71%为男性)年龄在20至75岁之间(中位数为61岁)的个体,他们是在年度健康检查期间招募的(扩展数据图1a)。排除了糖尿病患者,以避免高血糖的长期影响。因此,与大多数以前的关于糖尿病和肥胖的宏基因组学研究相比,我们的研究包括了相对健康的个体;体重指数(BMI)和糖化血红蛋白(HbA1c)的中位数(四分位数范围)分别为24.9 kg m−2(22.2–27.1 kg m−2)和5.8%(5.5–6.1%)(补充表1)。本研究分析的主要临床表型是胰岛素抵抗(IR),我们将其定义为胰岛素抵抗的稳态模型评估(HOMA-IR)得分至少为2.5(参考文献13)。我们还分析了粪便代谢物与代谢综合征(MetS),一种与IR相关的病理之间的关联。IR和MetS的临床特征在除了血压和性别比例之外的方面基本重叠,这两个方面在IR和正常胰岛素敏感性(IS)的个体之间没有差异(补充表1)。使用两种基于质谱(MS)的分析平台进行的无靶向代谢组学分析,分别鉴定了195种和100种注释的粪便和血浆亲水性代谢物,以及2654种和635种注释的粪便和血浆脂类代谢物(扩展数据图1a)。为了确定微生物功能、粪便代谢物和预测基因的总体差异,分别总结了共丰度组(CAGs)和KEGG类别。使用CAGE方法,获得了外周血单核细胞(PBMCs)的转录组信息,该方法可以在转录起始位点的分辨率上测量基因表达。
为了检验粪便样本的组学数据如何预测IR,我们首先比较了基于随机森林分类器的受试者工作特征(ROC)和曲线下面积(AUC)。模型的预测变量是使用最小冗余最大相关算法从粪便16S、代谢组、宏基因组和它们的合并数据集中选择的(补充表2)。我们发现,粪便代谢组学数据的选定特征在预测IR方面通常优于16S和宏基因组学的特征(图1a),这表明粪便代谢组学可以用来研究IR的发病机制。
01
粪便中的碳水化合物在IR中增加
接下来,我们研究了临床表型和粪便代谢物CAGs之间的关联(图1b和补充表3-8)。在相关性和回归分析中,对主要的混杂因素,即性别和年龄进行了调整。在亲水性代谢物中,与IR有显著关联的CAGs大多是碳水化合物代谢物,主要是单糖(亲水性CAGs 5、12和15;图1b,上)。已知作为碳水化合物发酵产物的短链脂肪酸(SCFAs)也在IR中增加(亲水性CAG 8)。亲水性CAG 仍未被注释,因为它包括了来自不同途径的代谢物(补充表5)。利用KEGG途径富集分析,发现这些与IR相关的亲水性CAGs中的代谢物确实参与了碳水化合物代谢(扩展数据图2a)。具体来说,我们发现主要的单糖,如果糖、半乳糖、甘露糖和木糖,与IR显著相关(图1c)。在SCFAs中,丙酸特别在IR中增加(扩展数据图2b),这与它在糖异生中的作用一致。粪便中的单糖在MetS、肥胖和糖尿病前期中也有类似的增加(图1d和扩展数据图2c,d)。相反,双糖则显示出弱的或没有关联(扩展数据图2b-d)。这些发现表明,碳水化合物降解的终产物,如单糖,它们可以被宿主直接吸收和利用,特别是在IR和MetS的个体的粪便中增加。支持这些发现的是,我们对来自Twins UK队列的先前发表的粪便代谢组学数据的分析显示,粪便中的单糖,尤其是葡萄糖和阿拉伯糖,与肥胖和HOMA-IR呈正相关,这两者都与IR相关(扩展数据图3a-c和补充表9)。类似地,在来自HMP2数据的没有炎症性肠病(IBD)的少数个体中,粪便中的果糖、葡萄糖和半乳糖的峰强度与BMI相关(扩展数据图3d)。这些发现表明,粪便中的碳水化合物在IR和相关病理中增加,这种改变在不同的人群中都一致地观察到。
除亲水性代谢物外,粪便脂质CAGs也与IR相关(图1b)。溶血磷脂、胆汁酸和酰基肉碱与IR和MetS相关,这与先前的报道相一致。其中,一个主要由双半乳糖/葡萄糖二酰基甘油(DGDG)(脂质CAG 11)组成的脂质CAG引起了作者的注意,因为据报道DGDG来源于细菌。这些脂质在其结构中含有葡萄糖和/或半乳糖,尽管它们在哺乳动物中的生物学功能大部分尚不清楚。这一类别中的大部分DGDGs与一些前体二酰基甘油和单糖(即葡萄糖和半乳糖)呈正相关(扩展数据图4a)。由于二酰基甘油在IR发病机制中起着重要作用,这类代谢产物的生物学功能尤为重要。值得注意的是,脂质CAG 41中具有不同酰基链的DGDGs与IR没有关联(附表7),这意味着脂质酰基链的差异可能具有先前报道的生理重要性。
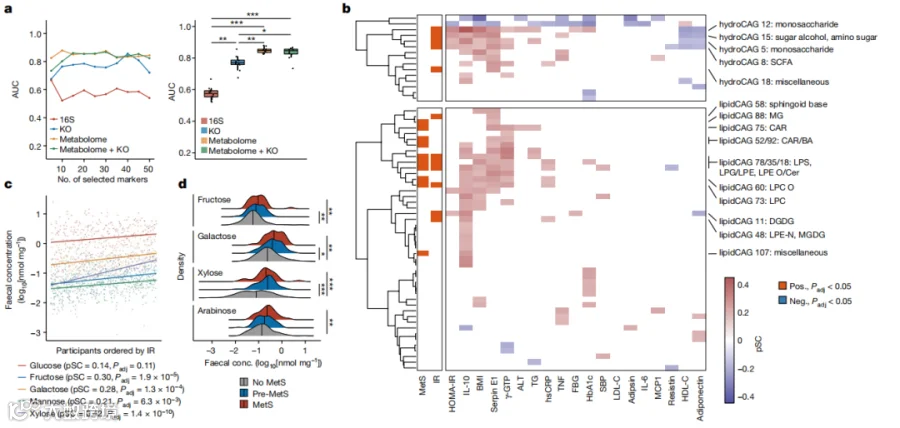
图1.粪便碳水化合物代谢物在IR中发生明显变化。a,左侧:使用随机森林分类器的AUC值,基于属水平16S数据(n=282)、KEGG orthologue(KO)水平宏基因组数据(n=266)、粪便代谢组和宏基因组(KEGG orthologue)+粪便代谢组数据(n=266)来预测胰岛素抵抗(IR)。在x轴上,选择的特征标记数量逐渐增加。右侧,箱线图显示所选特征获得的AUC值。b,粪便亲水代谢物(hydroCAG,顶部)和脂质代谢物(lipidCAG,底部)以及临床表型和标志物(n=282)。左侧的双列热图表示使用基于排名的线性回归分析的主要临床表型(IR和MetS)的相关性,而主热图显示了根据年龄和性别调整后的部分Spearman相关性(pSC)与代表性代谢标志物。只有Padj < 0.05的CAGs着色。c,HOMA-IR和粪便单糖水平之间的pSC。描述了系数(pSC)和Padj值(n=282)。d,MetS中的粪便单糖水平(n=306)。
02
IR中的微生物-代谢物关系
我们接下来研究了与IR相关的肠道菌群的改变及其功能。肠道菌群的多样性在个体之间有所不同(扩展数据图5a-e)。我们使用16S rRNA测序数据分析了研究参与者的属水平的微生物组成,鉴定出四个细菌群(扩展数据图5f)。群1以Lachnospiraceae科的细菌为主,如Blautia和Dorea,而群2以Bacteroidales为特征(如Bacteroides、Parabacteroides和Alistipes)和Faecalibacterium。群3包含了放线菌门的属。群4没有形成明显的网络。我们还可以根据他们的分类学特征,将研究参与者分为四个簇,A到D(图2a)。簇C的个体明显富含Bacteroidales的群2,而簇D的个体则显示出群1和3细菌的更高丰度(扩展数据图5g)。值得注意的是,IR的比例(图2a;P = 0.0071)在簇C中显著较低。与IR和MetS相关的其他代谢参数,如HOMA-IR、BMI、甘油三酯、高密度脂蛋白胆固醇(HDL-C)和脂联素,也在簇C(IR比例最低的簇)和其他三个簇之间有所不同(图2b和补充表10)。在富含群1和3细菌的个体中,IR的比例一致地高于富含群2细菌的个体,这是基于宏基因组测序数据鉴定的(扩展数据图5h)。HOMA-IR与Rikenellaceae科的Alistipes属以及来自Bacteroides、Bifidobacterium和Ruminococcus的几个物种呈负相关(扩展数据图5i和补充表11和12),部分重现了先前关于肥胖个体的报告。值得注意的是,不同的属和物种与其他临床标志相关,这表明微生物分类单元和临床表现之间的个体关联不如共丰度分析那样稳健。
接下来,我们根据显著的正相关或负相关(补充表13)构建了一个微生物-代谢物网络。尽管粪便中的SCFAs和脂类,如DGDG,与IR-和IS-相关的细菌群都有相关性,但IR-相关的粪便碳水化合物主要与群1和4中的属有相关性,其中最突出的是Lachnospiraceae科的Dorea(图2c,d)。相反,这些碳水化合物的大部分与群2细菌中的IS-相关的属,如Bacteroides、Alistipes和Flavonifractor呈负相关(图2d和扩展数据图5j),与群1中的细菌的相关性很小。因此,粪便中的碳水化合物水平在参与者簇之间有明显的差异(扩展数据图5k)。以前的研究表明,一些Lachnospiraceae科的物种参与了多糖的发酵,而Alistipes则在动物性饮食而不是富含多糖的饮食中增加。这些发现突出了碳水化合物降解产物和IR-和IS-相关细菌之间的紧密联系,表明这些细菌可能参与了IR中粪便碳水化合物谱的异常。
IR相关的粪便碳水化合物也与与碳水化合物代谢和运输相关的KEGG途径有关,如磷酸转移系统(PTS)、淀粉和蔗糖代谢、半乳糖代谢,而与与碳水化合物分解相关的途径呈负相关,如糖酵解和丙酮酸代谢(图2e和补充表14和15)。这些途径也与图2a中定义的参与者簇和图2d中定义的与碳水化合物相关的属有明显的相关性。氨基酸代谢也有所不同,特别是在簇C和D之间,而脂类代谢则没有与菌群显示出明显的关联(扩展数据图6a,b和补充表16)。尽管PTS和淀粉和蔗糖代谢等碳水化合物途径与HbA1c和γ-GTP呈强正相关,但与其他IR标志的相关性则相对稀疏(扩展数据图6c和补充表17),表明代谢物对临床表现的敏感性更高,如图1a所示。PTS是细菌将糖类作为能量来源转入自身的必要组成部分。对KEGG同源物的详细分析显示,粪便碳水化合物和参与者簇主要与与双糖和氨基糖相关的PTS有关(扩展数据图6d,e和补充表18),表明微生物通过PTS利用糖类的偏好可能影响代谢物的水平。水解寡糖和双糖的糖苷酶也与粪便单糖相关(扩展数据图6f)。细胞外葡萄糖苷酶,如β-果糖糖苷酶(K01193,KEGG同源物数据库)、淀粉蔗糖酶(K05341,KEGG同源物数据库)和寡糖-1,6-葡萄糖苷酶(K01182,KEGG同源物数据库),预测可以将蔗糖和糊精水解为葡萄糖和果糖(扩展数据图6g,h),显示出最高的正相关性,特别是与粪便葡萄糖。相反,与淀粉利用相关的葡萄糖苷酶,如α-淀粉酶(K01176和K07405,KEGG同源物数据库),则与粪便碳水化合物呈负相关。重要的是,这些糖苷酶基因的丰度在图2a中定义的参与者簇C和其他三个簇之间有显著差异,表明分类学特征在很大程度上解释了糖苷酶的变异(图2f,扩展数据图6h和补充表18)。一致地,在簇D中丰度较高的Blautia和Dorea的基因组中,主要保留了分解双糖的基因,而在簇C中丰度较高的Bacteroidales的基因组中,几乎缺乏这些基因(扩展数据图6i)。综上所述,我们的发现揭示了四个具有独特分类学特征和碳水化合物代谢的不同群体,它们以糖类的利用和降解为特征,与IR及其相关标志相关。
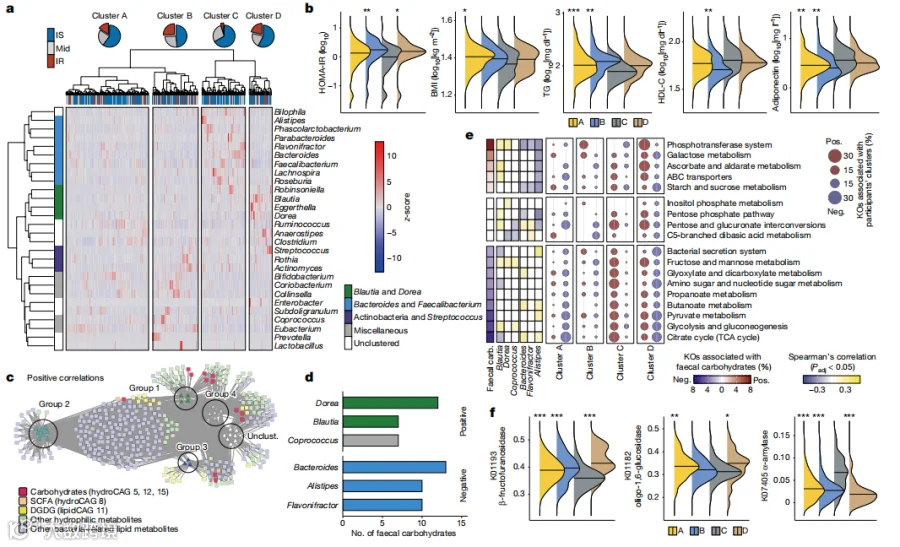
图2.IR相关的粪便代谢物与肠道微生物群和微生物遗传功能的改变有关。a,属水平细菌的共丰度聚类及其丰度(n=282)。参与者根据其分类学特征被分为A至D四个簇。显示了IR患者的比例。b,参与者簇中的HOMA-IR、BMI、三酰甘油(TG)和高密度脂蛋白胆固醇(HDL-C)水平。c,共现微生物群与粪便代谢物(n=282)之间的细菌-代谢物网络。包括所有粪便亲水性和与细菌相关的脂质代谢物。仅显示正向且显著(Padj < 0.05)的Spearman相关性。与图1b中的碳水化合物有关的CAGs中的代谢物以红色突出显示。Unclust.表示未聚类。d,菌属与粪便碳水化合物之间显著正相关和负相关的数量。显示了每个相关性中排名前五的属。e,与碳水化合物代谢和膜转运相关的KEGG通路、粪便碳水化合物、与粪便碳水化合物正相关或负相关的前三个属以及参与者簇。f,参与者簇中参与糖苷酶的代表性KEGG orthologues的丰度(n=266)。丰度经过平方根反正弦转换。
03
粪便碳水化合物和胰岛素抵抗中的炎症
与先前的报告一致,宿主的细胞因子、代谢组和转录组特征与IR高度相关(补充表19-21)。此外,这些PBMC基因在功能上涉及炎症(扩展数据图7a)并可能来自单核细胞(补充表21)。一些研究表明,微生物成分如脂多糖在促进代谢性疾病的炎症中有作用。然而,微生物代谢是否涉及低度炎症仍不清楚。因此,我们试图推断宿主IR的炎症特征与粪便碳水化合物之间的可能关联。首先,基于个体代谢物、细菌、转录本和细胞因子与IR相关的交叉组学相关性网络显示,粪便碳水化合物与细菌和宿主IR相关特征,特别是细胞因子,有着强烈的联系,表明这些代谢物是IR中宿主-微生物网络的枢纽(图3a,扩展数据图7b,c和补充表22)。差异丰度,计算为它们在IR和IS中的丰度比,最明显地表现在粪便碳水化合物和细胞因子之间的关联。值得注意的是,IL-10,一种血浆细胞因子,与粪便碳水化合物显示出最突出的关联,而与PBMC来源的转录本的关联适中,支持了最近的研究,显示其促进IR的悖论效应。粪便碳水化合物中等地解释了IL-10的方差,以及在较小程度上,脂联素、瘦素和serpin E1的方差,表明粪便碳水化合物特别与这些细胞因子相关(图3b)。虽然粪便碳水化合物解释的方差比例低于血浆代谢物,但比属级丰度高得多,突出了粪便代谢物在连接肠道微生物和宿主炎症反应中的作用。我们接下来试图推断通过因果中介分析这些细胞因子是否介导了粪便碳水化合物对宿主代谢的影响。我们发现,IL-10、serpin E1、脂联素和瘦素介导了粪便碳水化合物和宿主IR标志物如HOMA-IR之间的大多数体外因果关系(图3c,扩展数据图7d和补充表23)。值得注意的是,代谢物和细胞因子之间存在独特的对应关系;例如,IL-10介导了果糖、甘露糖、木糖和鼠李糖的效应,但不介导其他代谢物的效应。虽然这些独特对应关系的生物学意义有待研究,但IR中粪便微生物、代谢组和宿主炎症表型的联合分析揭示了一种以前未被认识的相互作用,即过量的单糖可能影响宿主细胞因子的表达。
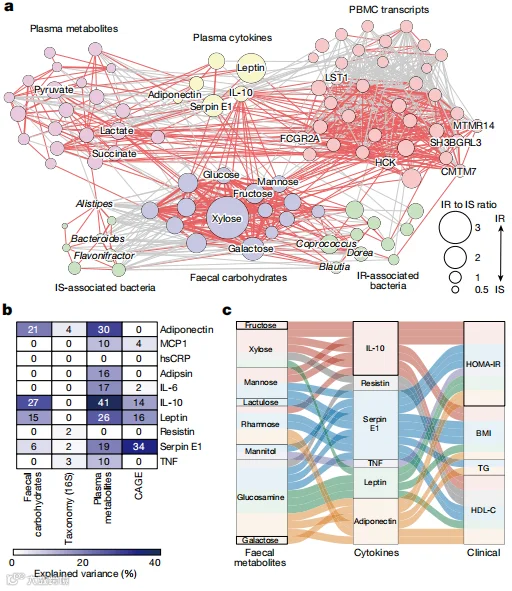
图3.粪便碳水化合物代谢物与IR中的细胞因子水平相关。a,基于IS(Insulin Sensitive,胰岛素敏感)、中等(即,HOMA-IR >1.6且<2.5)和IR(Insulin Resistant,胰岛素抵抗)样本的所有组学信息(n=46、70和275),构建了粪便碳水化合物代谢物(紫色)、粪便细菌(绿色)、血浆亲水性代谢物(粉色)、细胞因子(黄色)和PBMC基因(红色)之间的网络。分析包括与IR显著相关的宿主衍生标志物(见补充表19–21),图1b和扩展数据图5f中分别鉴定的15种粪便碳水化合物和20个属。为构建组学网络,计算了根据年龄、性别、BMI和FBG调整的配对pSC,并显示了Padj < 0.05的相互作用。线的宽度表示系数的绝对值,红色和灰色线分别表示正相关和负相关。b,使用随机森林分类器预测每个组学数据集预测的10种血浆细胞因子的解释方差。c,冲积图显示了血浆细胞因子显著介导了粪便碳水化合物对宿主代谢标志物的silico效应。线显示了中介效应,颜色表示由单个细胞因子介导的关联。
04
实验模型中的IS相关细菌
人类多组学分析的上述发现揭示了碳水化合物代谢物与IR病理之间的关联。为了探讨肠道菌群、粪便碳水化合物和代谢性疾病之间的因果关系,我们首先分析了22种人类粪便IS-和IR相关细菌的细菌培养物中的代谢物。这些细菌是根据属级共生(图2a,b)和种级(扩展数据图5i)特征的发现选择的。198种代谢物的主成分分析图表明,Bacteroidales,一种代表性的IS相关细菌目,在PC1上显示出不同的代谢特征(扩展数据图8a,b和补充表24)。导致组间分离的前10种代谢物包括几种氨基酸和发酵产物,如琥珀酸和富马酸,这些代谢物的大部分由Bacteroidales优先产生(扩展数据图8b,c)。我们在细菌培养物中检测到了与IR相关的15种碳水化合物中的13种(图1b)(扩展数据图8b)。这些碳水化合物中的大多数沿着PC1负向排列,表明这些代谢物与Bacteroidales呈负相关。葡萄糖、甘露糖和氨基葡萄糖被Bacteroidales与其他目相比优先消耗,而乳果糖主要由Eubacteriales产生(扩展数据图8d)。Alistipes indistinctus是最能消耗各种碳水化合物的细菌(扩展数据图8e,f)。这些发现表明,Bacteroidales物种是几种碳水化合物的强效消耗者,推动了它们的发酵产物的产生。
我们接下来测试了在人类队列发现中与IS相关的七种候选细菌的潜在治疗效果。在高脂饮食的小鼠中,给予A. indistinctus、Alistipes finegoldii和Bacteroides thetaiotaomicron的小鼠的餐后血糖水平特别降低(图4a)。胰岛素耐受试验也显示,这些菌株改善了IR,最明显的是A. indistinctus的给药(图4b,c)。A. indistinctus的给药改善了体重增加、肝脏的异位甘油三酯积累和葡萄糖耐受(扩展数据图9a-d)。A. indistinctus治疗的小鼠的血清高密度脂蛋白胆固醇(HDL-C)、脂联素水平也有所改善,甘油三酯水平也有所降低(扩展数据图9e-g)。高胰岛素血糖恒定钳分析的结果表明,A. indistinctus的给药显著改善了IR,特别是全身葡萄糖清除(扩展数据图9h-j)。A. indistinctus和A. finegoldii治疗的小鼠的肝脏和附睾脂肪中的AKT磷酸化增加(扩展数据图9k,l),表明肝脏和脂肪组织中的胰岛素信号转导有所改善。这些发现揭示了A. indistinctus给药在改善饮食诱导的肥胖和IR方面的潜力。
在机制上,代谢测量显示,A. indistinctus治疗的小鼠的碳水化合物氧化显著降低,表明碳水化合物的利用受到限制(扩展数据图9m和补充表25)。由于饮食摄入和运动活动保持不变(扩展数据图9n,o),我们推断A. indistinctus的治疗减少了肠道中宿主可利用的碳水化合物。在这方面,A. indistinctus的给药显著改变了盲肠代谢物,其特征是几种碳水化合物,包括果糖,一种脂肪合成的单糖(扩展数据图10a-c和补充表26)的减少。果糖在血清中也有类似的降低(扩展数据图10d)。重要的是,胰岛素耐受试验的AUC与盲肠单糖果糖、葡萄糖和甘露糖呈正相关(图4d)。综上所述,这些发现表明,A. indistinctus改善了小鼠的IR,并影响了肠道碳水化合物代谢物,支持了我们在人类队列中的观察。
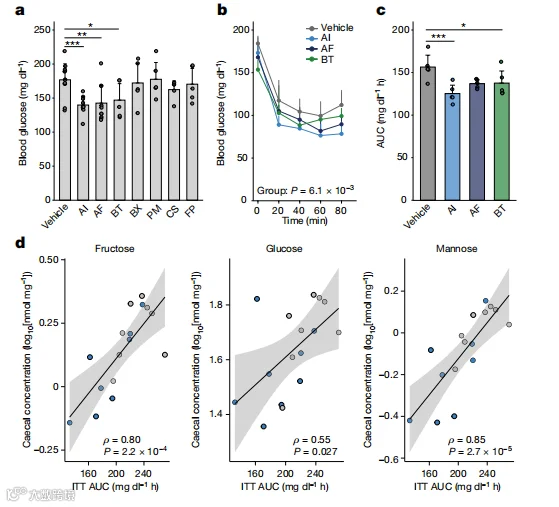
图4.在实验模型中,IS相关细菌改善了IR。a,饲喂高脂饮食的小鼠在细菌给药后4周的餐后血糖。n=12(对照组),n=10(A. indistinctus和A. finegoldii组),n=5(其他组)只小鼠。b和c,胰岛素耐量试验期间的血糖水平(b)和AUC(c)(每组n=5)。d,胰岛素耐量试验的AUC与A. indistinctus(蓝色)或对照(灰色)组盲肠中果糖、葡萄糖和甘露糖水平之间的相关性。
讨 论
为了深化我们对IR中宿主-微生物关系的理解,我们使用多模态技术进行了一项全面而广泛的研究,探讨了人类肠道微生物组和代谢性疾病之间的相互作用。虽然肠道微生物对碳水化合物的代谢已经被认为影响肥胖和糖尿病前期的发病机制,但由于缺乏详细的代谢组信息,人类中的实际机制联系一直难以捉摸。在这方面,我们方法的主要优势是,我们结合了粪便代谢组学,记录了超过2800种注释的代谢物,以及微生物组和宿主病理学。这种基于代谢组的方法使我们能够识别与IR相关的粪便代谢物,发现粪便碳水化合物与IR的低度炎症之间的关联,以及在实验条件下有效地选择候选菌株进行功能验证(扩展数据图10e)。综上所述,我们的研究突出了全面的组学策略在探索微生物代谢及其产物在IR发病机制中的作用的优势。过量的单糖有可能促进异位脂质积累,同时激活免疫细胞,导致低度炎症和IR。果糖是一种广为人知的炎症和IR的危险因素,因为它在脂质积累中起着作用,而半乳糖已被证明参与激活的免疫细胞的能量代谢。我们的体内研究证实,A. indistinctus的给药改善了脂质积累和IR,同时降低了肠道单糖水平(图4d)。然而,我们意识到需要进一步的机制研究来检查吸收的动力学及其对宿主代谢的影响。特别是,Alistipes菌株如何抑制碳水化合物代谢是一个有趣的问题(例如,这些细菌本身是否抑制碳水化合物代谢,或者它们是否与其他共生菌相互作用),因为它将直接开启一种新的治疗策略的可能性。考虑到A. indistinctus改善了全身IS(扩展数据图9i),研究肝脏以及包括骨骼肌和脂肪组织在内的周围组织中的胰岛素信号转导的参与是很重要的,以及这些组织中特定脂质分子(如神经酰胺和二酰甘油)的积累。这些研究有可能揭示导致A. indistinctus介导的IR改善的潜在机制。最后,人类研究中有两名参与者无法在早上收集他们的粪便,这可能会由于缺乏对一天中时间和禁食条件的严格控制而影响结果。因此,我们认为需要进行纵向研究,及时记录饮食习惯,以在考虑潜在的混杂因素的同时,剖析微生物代谢对糖尿病及其并发症轨迹的复杂影响。
本文译自:Takeuchi T, Kubota T, Nakanishi Y et al. Gut microbial carbohydrate metabolism contributes to insulin resistance. Nature. 2023 Sep; 621(7978):389-395.
