
奇辉观点
肠道微生物群通过多种机制深刻影响能量平衡,这一领域日益增长的知识为体重管理提供了新机会。饮食、身体活动是影响肠道微生物组的体重管理的关键因素,微生物组可能为干预效果和长期体重维持提供新的生物标志物。合理操纵微生物组可以提高现有疗法的效果并可能产生新的治疗方法,为医生和患者提供新的体重管理选择。要实现这一前景,需要克服几个基本挑战,包括应对肠道微生物组的生态敏感性、个体间变异性、发展可塑性和肠道微生物组对人类生理学的多效性影响。人类需要增加对肠道微生物组功能和产品的关注度,以便有效地利用微生物群来改善全球代谢健康。
论文ID
本文译自:Rachel N Carmody , Jordan E Bisanz. Roles of the gut microbiome in weight management. Nat Rev Microbiol . 2023 Aug;21(8):535-550. doi: 10.1038/ s41579-023-00888-0.
发表杂志:Nat Rev Microbiol
影响因子:68.8
通讯作者:Rachel N. Carmody
作者单位:Harvard University
摘要
超重、肥胖、营养不良及其各自的后遗症对全球个人和公共健康造成了毁灭性的影响。传统的治疗方法,包括饮食、运动、药物和/或手术,已经显示出不同程度的成功,这创造了对具有长期效果的新解决方案的迫切需求。由于测序、生物信息学和无菌实验的变革性进步,我们现在了解到肠道微生物群通过多种机制深刻影响能量平衡,这些机制影响能量平衡方程的两边。我们对微生物对能量代谢贡献的日益增长的知识强调了体重管理的新机会,包括微生物群意识的现有工具改进和针对微生物群的新疗法。在这篇综述中,我们综合了当前关于肠道微生物群与现有体重管理策略之间双向影响的知识,包括基于行为和临床的方法,并结合了一项主题级别的元分析,对比了体重管理策略对微生物群组成的影响。我们考虑了对肠道微生物群的新兴理解如何改变我们对体重管理的前景,以及为了使以微生物群为重点的解决方案取得成功必须克服的挑战。
正文
慢性能量不平衡影响了全球三分之一的人口。根据世界卫生组织最近的估计,全球有19亿成年人和3.8亿儿童超重或肥胖,自1975年以来肥胖率增长了两倍。另外还有4.62亿成年人和2亿儿童营养不良,营养不良导致5岁以下儿童死亡的45%。发展中国家和低社会经济地位的个体面临不成比例的营养不良负担,并且是超重和肥胖增长速度最快的一些地区。相关的疾病可以同时降低生产力并增加医疗费用,加强了贫困和不良健康之间的联系。因此,能量不平衡的后果对个人和公共卫生、经济发展和社会正义具有深远和持久的影响。
这个问题规模庞大且不断增长的一个原因是现有的治疗显示出有限的长期成功。超重和肥胖通常通过生活方式干预来治疗,这些干预通过降低卡路里摄入和增加体力活动来诱发负能量平衡,肥胖还通过药物和/或手术进行治疗。这些干预通常在短期内成功,大多数超重的人能够在6个月内减去超过初始体重的5%。然而,几乎总是会出现体重反弹,因为急性负能量平衡触发了促进资源节约和降低静息代谢率的代谢适应。营养不良通常用即食治疗食品来治疗,这些食品旨在增加卡路里和/或蛋白质摄入,使用抗生素来对抗共感染,以及改善食品安全的结构变化。然而,这些干预通常证明不足以纠正营养不良,特别是在幼儿中。使问题更加复杂的是,早期生活中的营养不良可以改变发育,然后使个体在成年后容易患上代谢性疾病,一些影响甚至可以传递给后代。
仍然迫切需要新的体重管理方法。最近的变革性研究揭示了肠道微生物群对人类生理学的深远和广泛影响,包括能量平衡。肠道微生物群已被证明对体重管理的现有工具敏感,包括饮食和运动、药物和手术干预,如胃旁路手术。关键的是,肠道微生物群的变化也可以调节干预措施的效果,这表明合理操纵肠道微生物群可能有助于体重管理。在这里,我们概述了肠道微生物群在能量代谢中的作用,回顾了肠道微生物群与现有体重管理工具之间的双向影响,并评估了在发展针对能量平衡的微生物群导向治疗方面的机会和挑战(图1)。尽管迄今为止的大多数研究都集中在肠道微生物对超重和肥胖的贡献上,但只要可能,我们就会提请注意阐明和操纵肠道微生物群在营养不良中的作用的并行进展,这是微生物群靶向医学的一个有希望的新领域。
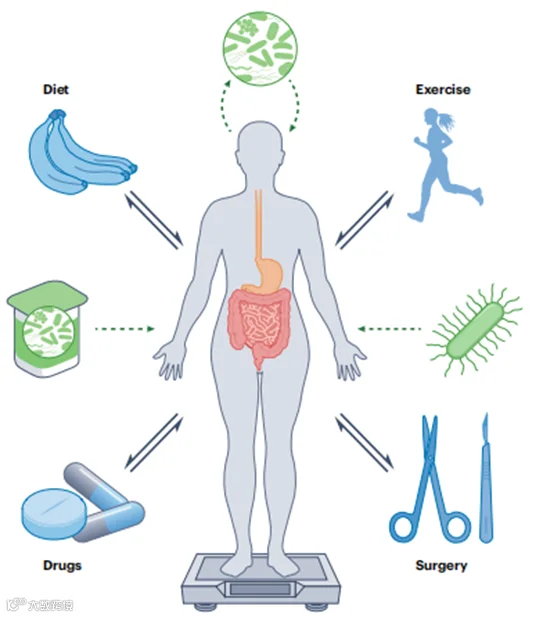
图1 | 肠道微生物群与关键的生活方式和临床体重管理方法之间的相互影响。常见的调节体重的干预措施(蓝色),如饮食、运动、药物和手术,影响肠道微生物的结构和功能,而这些肠道微生物群的变化反过来又改变了干预措施的效果。肠道微生物对体重管理的贡献通过新兴的微生物群导向治疗(绿色)被针对,包括设计用于支持有益微生物的植入或生长的食品,体重减轻后的自体粪便微生物群移植和下一代益生菌。
肠道微生物群与能量代谢
在无菌条件下饲养的无菌小鼠与常规饲养的小鼠或曾经无菌后被植入小鼠或人类肠道微生物群的小鼠相比,具有较低的脂肪率,尽管无菌动物吃得更多且消耗的能量更少。在携带抗生素消除肠道微生物群的小鼠中也观察到了类似的结果。这些研究表明,微生物群落对宿主能量状态有强烈的、通常是净正面的影响。此外,许多研究表明,通过微生物群移植,可以在无菌动物中重现肥胖、胰岛素抵抗、低度炎症和/或提高的生热等代谢表型,表明肠道微生物群在能量代谢中有因果影响。与常规小鼠相比,无菌动物在生理和免疫学上存在异常,包括与能量平衡相关的表型,如肠道屏障功能、参与营养获取和利用的酶的表达以及吸收表面的粗略生理学。人到小鼠的微生物群移植也不完全复制人类供体组成,而无菌动物的资源密集性质常常导致实验能力不足。尽管存在这些限制,无菌动物模型仍然是我们建立微生物群组成和宿主能量平衡之间因果关系的最好工具之一。
无菌研究表明,肠道微生物群在能量平衡方程的两边都有因果调节作用。例如,肠道微生物群通过短链脂肪酸(SCFA)介导的激素分泌以及直接微生物合成与肠神经系统和中枢神经系统相互作用以调节饥饿和饱腹感的神经递质和激素模拟物来调节宿主能量摄入(框1)。肠道微生物群通过增强小肠脂质吸收以及通过发酵将小肠中未消化的碳水化合物和蛋白质转化为具有残余热量价值的SCFA和其他代谢产物,从而增加饮食能量的收获(图2)。肠道微生物群通过生成可供选择的宿主组织使用的代谢底物、调节胆汁酸代谢以及影响控制脂肪酸吸收、脂肪分解和生热的宿主基因表达来指导能量利用(图3)。最后,肠道微生物群通过在发育期间训练系统免疫反应性、影响肠道屏障完整性以及产生具有可变代谢健康后果的促炎产物(如脂多糖和鞭毛蛋白)来影响能量平衡和炎症之间的相互作用(框2)。由于这些观察结果许多来自动物研究,未来的人类研究将需要确立其转化相关性。
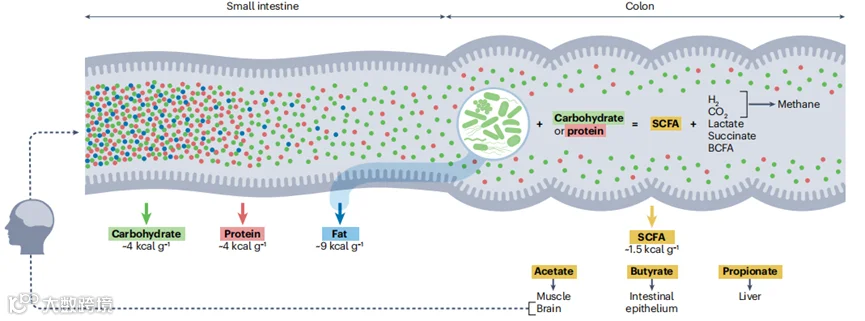
图2 | 肠道微生物组增强饮食能量收获。宿主酶可分解的宏观营养素在小肠中被消化。小肠吸收的宏观营养素提供了通过生物化学可预测的能量(碳水化合物,约4千卡/克;蛋白质,约4千卡/克;脂肪,约9千卡/克)。饮食脂肪在小肠近端很容易被吸收,尽管脂肪消化传统上完全依赖宿主酶,但在动物模型中肠道微生物组对小肠脂质吸收的贡献以及宿主-微生物在脂质乳化中相互作用挑战了这一观点。相比之下,普遍接受的是微生物增强了碳水化合物和蛋白质的消化。小肠消化的碳水化合物和蛋白质的比例随着宏观营养素的结构形式(例如,糖比纤维高)、餐食组成(例如,纤维少的比纤维多的高)、热处理(例如,煮熟食物高)和物理处理(例如,更小粒度的食物高)而变化。逃过小肠消化的营养物质会被结肠肠道微生物群发酵,产生一系列具有能量影响的代谢产物。肠道微生物组从饮食中的缬氨酸、亮氨酸和异亮氨酸产生支链脂肪酸(BCFAs),以及其他有机酸如乳酸和琥珀酸。然而,未消化的碳水化合物是微生物发酵的主要燃料,从中肠道微生物组生成短链脂肪酸(SCFAs)乙酸、丁酸和丙酸。这些SCFAs被宿主吸收,并在多种组织中有助于能量代谢,乙酸支持肌肉和大脑,丁酸供应结肠上皮细胞高达60-70%的能量需求,丙酸用于肝脏的糖异生。SCFAs的能量回收被估计为约1.5千卡/克(参考文献157),不到小肠消化的碳水化合物的一半。因此,当营养物质直接被消化而不是发酵时,宿主收获的能量更多。尽管如此,SCFAs在工业化人群中占每日能量需求的约5-10%,在饮食加工最少和/或富含纤维的人群中几乎肯定占有更大的比例。尽管SCFAs长期以来主要被赞赏为能量回收的工具,但最近的研究显示SCFAs具有强大的信号功能,可以调节能量摄入(虚线箭头;另见框1)、能量利用(图3)和炎症(框2)。这些多效性效应有助于解释为什么有关高脂饮食有无SCFA补充的研究报告了SCFA对体重增加的抑制作用。宿主代谢产物,包括胆汁酸(图3)和免疫因子(框2),也与肠道微生物组双向相互作用,并影响其对能量平衡的贡献。
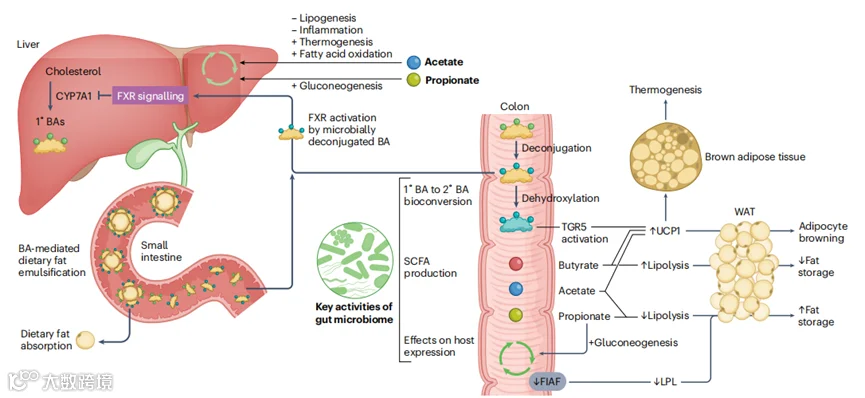
图3 | 肠道微生物组调节能量利用。来自肠道微生物组的信号作用于脂肪细胞,指导可用卡路里是分配到储存还是产热。早期的动物实验表明,肠道微生物群落的定植通过抑制肠道中的循环脂蛋白脂肪酶(LPL)抑制剂——禁食诱导的脂肪细胞因子(FIAF),导致脂肪细胞中脂肪酸的大量吸收,从而增加了脂肪细胞中的甘油三酯储存。短链脂肪酸(SCFAs)也影响脂肪、肠道和肝脏组织代谢状态。虽然由于跨研究结果和设计不一致,将SCFA产生与组织状态明确联系起来存在挑战,但许多研究发现乙酸、丁酸和丙酸具有不同的效应。例如,在储存能量的白色脂肪组织(WAT)中,乙酸和丙酸抑制了脂肪分解并促进了脂肪积累,而丁酸促进了脂肪分解和脂肪酸氧化。丁酸在消耗能量的棕色脂肪组织中增强了产热,而乙酸和富含乙酸的SCFA混合物增加了WAT的棕色化。丙酸在肝脏和肠道中促进了糖异生,而肝脏中的乙酸被报道可以减少脂肪生成,限制脂肪积累,增加产热和脂肪酸氧化基因的表达,并减少炎症。此外,特定的SCFA化合物可以在不同组织中以相反的方式影响生理学,正如乙酸或丁酸补充在脂肪组织中增加GPR43表达,而在结肠中减少它。这种不同的效应提出了通过输送特定SCFA来操纵代谢状态的可能性。肠道微生物对胆汁酸(BAs)的转化也调节能量代谢。肠道微生物通过脱结合和脱羟基将宿主产生的初级胆汁酸(1° BAs)转化为二级胆汁酸(2° BAs)。肠道微生物转化的BAs具有多样的信号功能。例如,脱结合的初级和二级BAs是核法尼酰X受体(FXR)的主要配体,FXR是一个转录因子,在BAs、胆固醇、脂质和葡萄糖代谢中具有关键的调节作用170。通过FXR信号,微生物对初级BAs的脱结合可以减少肝脏中CYP7A1的表达,CYP7A1是初级BA合成中的限速酶,可能对小肠中脂质吸收产生下游效应。此外,肠道微生物BA代谢在调节产热中起着关键作用。值得注意的是,二级BA石胆酸是G蛋白偶联受体TGR5的高亲和力激动剂,通过上调解偶联蛋白1(UCP1)诱导WAT和棕色脂肪组织产热。
肠道微生物群对能量状态的操纵是有益还是有害,当然取决于具体情况。在能量平衡的正负极端状态下的研究通常报告说,肠道微生物群加剧了宿主的能量表型。例如,与肥胖宿主相关的肠道微生物群具有结构和功能上的变化,这些变化增加了饮食能量收获的能力。在正能量平衡的动态状态下,包括在热量限制后体重反弹,在体重循环后再次引入致肥条件的复发性肥胖,以及在妊娠第三期的女性中,也发现了类似的促进体重增加的结果。肠道微生物群也可以加剧急性负能量平衡状态。例如,患有夸希奥科尔病(一种严重急性营养不良形式)的儿童的肠道微生物群表现出一种幼态状态,损害了营养吸收。同样,在体重迅速减轻的时期,如Roux-en-Y胃旁路(RYGB)手术后的肠道微生物群,已被证明在因果上有助于体重减轻。同样,接受非常低热量饮食(约每天800千卡)条件下的微生物群的无菌受体与接受饮食前微生物群的受体相比,脂肪率降低。关键的是,接受与夸希奥科尔病相关和RYGB后相关微生物群的无菌移植受体与无菌小鼠相比,体重和脂肪率较低,为具有对宿主能量状态净负面影响的微生物群提供了罕见的例子。
然而,肠道微生物群并不总是加剧能量不平衡。实际上,在至少某些条件下,宿主-微生物系统表现出一种动态能量缓冲的形式,在这种形式中,宿主短期减少能量摄取促进了一种微生物群,这种微生物群对能量状态的贡献增强。例如,在喂食营养匹配的生食和熟食饮食的小鼠中,已知这两种饮食在回肠消化性方面存在差异,较低的消化性生食饮食总体上导致体重减轻,但促进了一种肠道微生物群,这种微生物群本身通过提高接受生食饮食条件的微生物群的无菌受体的体重和脂肪率,促进了宿主能量状态的增加。肠道微生物群加剧与缓冲宿主能量状态的具体条件尚不清楚,但构成了研究的高优先级领域,因为这些条件揭示了肠道微生物群治疗操纵的杠杆。
将微生物特征与代谢表型联系起来的另一个挑战是,可能存在过去微生物群状态的当前后果。在小鼠和人类的研究中表明,通过脉冲治疗或慢性亚治疗剂量的抗生素破坏早期生活中的肠道微生物群,增加了成年期肥胖的风险。即使微生物群特征最终反弹,变得与对照组无法区分,也发现了破坏早期生活中肠道微生物群的代谢后果,这表明一些微生物群的代谢贡献将难以追踪。将破坏的早期生活微生物群与成年宿主能量表型联系起来的近因机制尚待阐明。例如,目前尚不清楚导致肥胖的过量能量是来自增加的食物摄入、减少的活动、降低的静息代谢率,还是减少到免疫或生殖的分配。同样,小鼠和人类的研究表明,肠道微生物群延迟成熟的信号可以先于婴儿营养不良的发生。什么损害了肠道微生物群的成熟尚不清楚,但有人提出分解隔室化和小肠细菌过度生长可能是促成因素。
值得注意的是,肠道微生物群的代谢编程甚至可能在出生前就开始,正如最近在小鼠中的一项研究所展示的,来自母体微生物群的SCFAs穿过胎盘屏障,在发育中的胚胎中与GPR41和GPR43受体结合,影响下游组织发育。由于无菌状态、抗生素处理或低纤维饮食导致SCFA生产不足的母体微生物群所生的小鼠,在成年后遇到高脂饮食时,患代谢综合征的风险比SCFA生产微生物群的母体所生的小鼠更高。关键的是,SCFA缺乏和SCFA生产的母亲所生的小鼠都被外科手术分娩并交叉寄养,因此这种差异不是由于垂直遗传SCFA生产微生物群。相反,是胚胎暴露于母体微生物群的SCFA决定了发育命运和未来饮食与代谢表型的相互作用。同样,最近的数据暗示胎儿暴露于微生物或垂直遗传的扰乱母体微生物群在塑造免疫发育方面的作用。尽管这些效应在人类中尚未被研究,但这些小鼠数据表明,微生物群可能是跨代调节体重管理干预措施效果的载体。
肠道微生物对能量代谢的影响可以在整个生命周期中投下阴影,并影响肠道之外的器官(图4)。这些多样化机制中固有的多效性有助于解释为什么很难预测肠道微生物扰动对宿主代谢反应的先验效应。
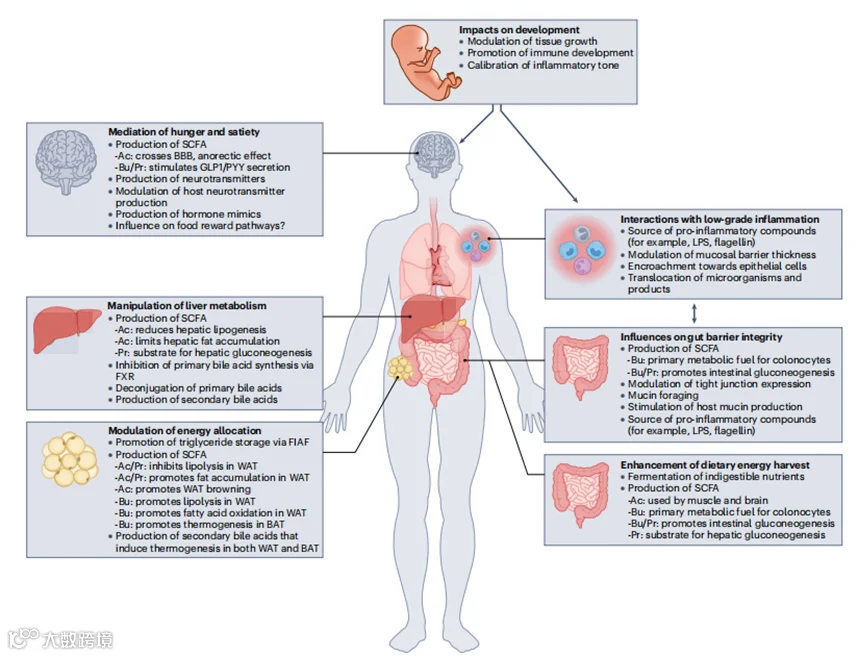
图4 | 肠道微生物对宿主能量状态影响的机制。肠道微生物组通过塑造代谢和免疫生理的早期影响,以及在整个生命过程中对饥饿和饱腹感、饮食能量收获、胆汁酸代谢和可用能量分配到储存与产热的动态影响,从而有助于宿主的能量代谢(框1和图2、3)。此外,肠道微生物组通过其在早期生活中对炎症基调的校准、促炎化合物的产生和对肠道屏障完整性的调节,调节能量代谢与低度炎症之间的相互作用(框2)。Ac,乙酸;BAT,棕色脂肪组织;BBB,血脑屏障;Bu,丁酸;FIAF,禁食诱导的脂肪细胞因子;FXR,法尼酰X受体;GLP1,胰高血糖素样肽1;LPS,脂多糖;Pr,丙酸;PYY,肽YY;SCFA,短链脂肪酸;WAT,白色脂肪组织。
塑造肠道微生物对体重管理贡献的饮食属性
体重管理饮食经常操纵饮食的宏观营养素含量,如低碳水化合物、低脂肪或高蛋白计划所示范。这些饮食之间可能存在一种错误的三元分法,因为饮食组成是一个零和游戏,一个比例的减少必然需要另一个比例的上升。例如,许多研究集中在饮食脂肪作为微生物结果的驱动因素,而实际上,这些饮食在糖和/或纤维含量以及整体食物与半纯化状态方面也有所不同。专注于给定宏观营养素的潜在陷阱由最近的报告突出显示,即对高脂肪生酮饮食和高脂肪非生酮饮食的微生物反应是不同的,生酮饮食导致双歧杆菌的丧失,可能由于酮类的抗菌效应。此外,常见的体重管理饮食通常不是等热量的,而是推动减少热量摄入,这本身就可以引起类似的临床结果和微生物反应(补充表1)。
然而,一些特定的饮食属性因其对肠道微生物组的影响而受到关注。膳食纤维向结肠提供可发酵的基质,丰富了合成碳水化合物活性酶的微生物,并上调SCFA的产生。实际上,长期低纤维摄入导致这些类群在小鼠中灭绝。在最近的人类研究中,高纤维饮食的消费增加了微生物组衍生的糖苷水解酶或已知的发酵类群,尽管对SCFA产生、微生物群多样性和宿主表型的影响各不相同。膳食纤维作为药物治疗的辅助手段,与阿卡波糖(一种用于2型糖尿病的α-葡萄糖苷酶抑制剂)一起使用,促进了SCFA产生微生物的生长,并降低了HbA1c水平,这是血糖的生物标志物。然而,纤维补充的效果并不总是有益的。例如,阿拉伯木聚糖或长链菊粉,这些在常见的非处方减肥补充剂中发现的分离纤维,对肠道微生物和宿主表型有不同的影响,阿拉伯木聚糖降低了胆固醇,菊粉促进了双歧杆菌的生长,但高菊粉剂量(每天30克)引发炎症和肝酶升高。分离的纤维补充剂通常针对双歧杆菌和乳酸菌的生长,因此可能无法复制富含多种多糖的饮食的效果。然而,由于可发酵纤维在刺激SCFA产生方面的能力不同,且SCFA在能量代谢的远端部位(图3和4及框1)的生物学效应不同,精确的纤维干预可能被用来设计SCFA池的组成,以改变能量代谢。
乳化剂和非热量人工甜味剂(NAS)已经成为工业化饮食中无处不在的添加剂。尽管通常被认为是安全的,但最近的研究表明这些化合物影响肠道微生物组及其对能量代谢的贡献。像羧甲基纤维素和聚山梨醇酯这样的乳化剂被观察到破坏肠粘膜,导致微生物群侵入、低度炎症、肥胖和通过微生物群移植传播的高血糖水平;然而,相对较少的充分动力的研究可用于确认这些发现在人类中。同样,在动物和人类的研究中表明,像糖精和三氯蔗糖这样的NAS化合物促进了葡萄糖不耐受,剂量在可接受的每日摄入量范围内导致微生物群传播的体重增加、代谢异常和炎症。然而,这些结果在后续的动物实验或涉及糖精和三氯蔗糖的人类研究中没有得到证实。这种不同组之间的可重复性缺乏和小型人类研究样本大小(每个干预组<20名参与者)表明,需要进一步调查和足够动力的双盲、安慰剂对照研究,以确定NAS的任何有害效应。
发酵食品和益生菌经常被认为是促进代谢健康的,但实证证据有限。由于营养成分和制备方法的差异,以及使用的不同种类和菌株组成,不同发酵食品和益生菌干预研究之间的结果直接比较通常很困难。最近的一项研究表明,富含各种发酵食品的饮食——例如奶酪、克菲尔、酸奶和康普茶——增加了肠道微生物组的多样性,并减少了血清中的促炎和抗炎细胞因子。然而,由于缺乏饮食标准化,目前尚不清楚哪些功能性食品及其相应的微生物负责这些效果。传统的基于乳酸菌的益生菌的免疫调节能力得到了充分记录,并且有越来越多的证据表明对葡萄糖稳态有益。然而,它们在体重调节方面的证据是有限的,高质量的研究很少。一项系统综述发现,酸奶消费与较低的体重、体重增加、身体质量指数、腰围和体脂相关,但由于混杂变量,因果关系仍不清楚。在肥胖人群中进行的最近一项随机、双盲、安慰剂对照研究中,用从泡菜中分离的益生菌Lactobacillus sakei治疗12周,导致体脂和腰围减少,但对体重或身体质量指数没有影响。尽管产品营销强劲,据我们所知,目前没有高质量的证据将康普茶与人类的免疫或代谢益处联系起来。
特定饮食与肠道微生物对体重管理的贡献
饮食是一个复杂变量,其多个维度影响微生物组的组成,包括热量含量、宏观营养素和微量营养素负荷、食物的准备和进食时间。尽管在受控的人类或动物研究中,每个维度都可以单独研究,但体重管理饮食通常结合了这些变量的组合,使得难以解读它们各自的贡献。这进一步因跨研究直接比较微生物组数据的困难以及跨个体比较微生物组特征的困难而变得复杂。为了量化体重管理干预措施对人类肠道微生物群的高层级效应,我们使用先前发表的方法比较了14项纵向研究中个体内部微生物群的变化,这些研究涉及常见的体重管理干预措施,包括热量限制、营养素调节、运动和RYGB手术(补充方法)。研究的选择基于设计和数据可用性(补充表2)。与以前的报告一致,我们发现α-多样性既不是健壮的也不是可重复的干预指标。然而,在14项研究中的12项中,体重管理干预在参与者间可重复地改变了肠道微生物群的组成(图5a)。
在解释这一结果时需要考虑的一个重要因素是,体重管理干预的效果不太可能超过个体间变异的影响。尽管并不总是报告,我们确定干预解释了肠道微生物群组成变化的0.6-9.0%,而个体间变异解释了55.2-87.0%,这取决于研究设计和距离度量。
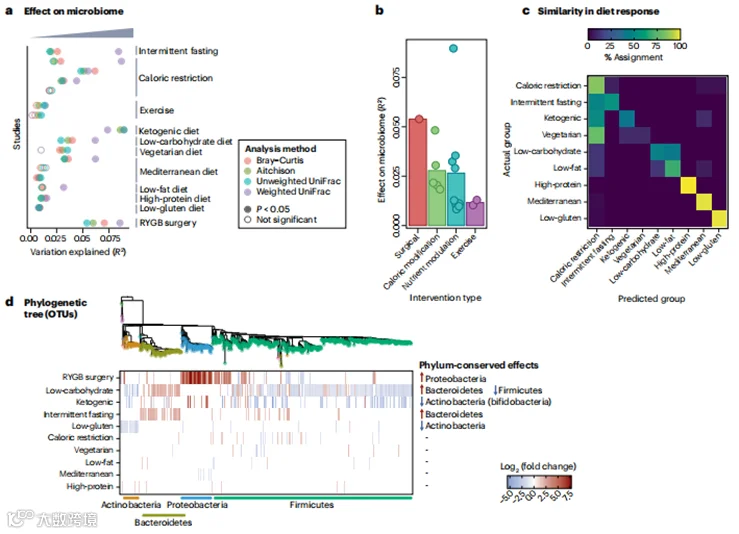
图5 | 体重管理干预措施对肠道微生物组成影响的元分析。
a,各种干预措施对肠道微生物组成的影响大小不一,通过在每项研究中归因于干预的肠道微生物组成变化的比例来衡量,这是一个不受跨研究微生物组成差异测量方式影响的稳健指标。该图示展示了几种最常用的距离度量方法,用以表征样本间差异,包括Bray-Curtis不相似性、Aitchison距离、未加权UniFrac距离和加权UniFrac距离。不同距离度量方法得出的结果差异反映了它们在处理相对丰度和系统发育关系方面的不同处理方式。
b,手术干预和饮食干预相比运动更能显著调节微生物组,通过解释变化的比例来衡量。该图示展示了对未加权UniFrac距离的分析,但使用其他常用距离度量方法也观察到类似结果。热量调整饮食包括间歇性禁食和旨在减少热量摄入的饮食,而营养素调节饮食包括涉及故意重新平衡宏观营养素的饮食(例如,低碳水化合物、低脂肪、高蛋白或生酮饮食)和地中海饮食。
c,为了确定肠道微生物对各种饮食的反应相似性,对数据拟合了一个机器学习模型。该模型显示,高蛋白、地中海和低麸质饮食对肠道微生物组有独特且容易区分的影响(由黄色方框指示)。相比之下,间歇性禁食、生酮和素食饮食的效果难以与一般热量限制区分开来,低碳水化合物和低脂肪饮食的效果也难以相互区分(由蓝绿色方框指示)。
d,对体重管理干预影响的有机体(操作分类单位OTUs)进行分析,并按其系统发育相关性水平排序,揭示了饮食在门水平上的保守效应。特定的门水平关联列在图表右侧。RYGB,Roux-en-Y胃旁路手术。参与此元分析的研究详情见补充表2和补充方法。
在我们的数据集中,干预持续时间跨度很大,从3天到12周不等。值得注意的是,无论距离度量如何,我们没有检测到效应大小和干预持续时间之间的显著相关性(rho = -0.09至0.2,所有P > 0.43,Spearman相关性),这加强了即使是短期的生活方式干预也可以重新编程肠道微生物组的观点。跨研究比较效应大小显示,RYGB手术对微生物群组成有最大的影响,其次是热量限制、营养素调节,最后是运动(图5b)。这些分类必然是不完美的,因为RYGB手术在一定程度上针对的是热量限制,营养素调节可能影响热量负荷,运动通常改变饮食。饮食干预的子分析显示,间歇性禁食、长期热量限制和生酮饮食对肠道微生物群组成有最大的影响,被识别为素食或低麸质的饮食影响最小(图5a)。为了理解哪些饮食引起了类似的效应,我们使用机器学习方法从饮食引起的肠道微生物群组成变化中预测饮食类型(补充方法)。我们发现该模型难以区分间歇性禁食、生酮饮食和素食饮食与热量限制,而高蛋白、地中海和低麸质饮食则容易区分(图5c)。对响应干预的微生物的系统发育检查(图5d)显示了高水平的信号,例如RYGB手术后的Proteobacteria增加或间歇性禁食或低碳水化合物饮食后的Bacteroides spp.增加,但也有饮食内和饮食间的特异性。尽管比较应谨慎解释,因为任何给定干预的可用研究数量很少,并且测序的性质没有捕捉到微生物群绝对丰度的降低,正如先前对极低热量饮食所确定的,但这些观察结果提供了对体重管理干预措施的比较性肠道微生物效应的初步了解。
饮食结果
将微生物组科学转化为实际应用的一个巨大机会在于使用微生物组作为预测工具。个体对减重干预措施的短期和长期反应的高度差异与各种行为和生物指标有关,但微生物组可能为干预效果和长期体重维持提供新的生物标志物。在小鼠中的原理验证数据表明,利用微生物群组成进行预测的分类模型在“悠悠球”式节食模式中以高准确率预测了体重重新增加。特定的微生物类群也与饮食效果有关。例如,Prevotella spp.的基线水平预测了在高纤维、全谷物饮食上的6周减重,而肠道微生物群多样性较高以及诸如Ruminococcaceae spp.和Lachnospiraceae spp.等类群的相对丰度预测了在TwinsUK队列中的健康女性在10年内体重增加。在最近的一项研究中,确定了在减重干预期间体脂变化的约22-38%可以通过基线微生物群来解释,某些Clostridia和Parabacteroides spp.表明了增加的损失。有趣的是,较高的基线微生物群多样性与较低的脂肪损失相关,这加强了追求高肠道微生物群多样性不太可能普遍有益的观点。当微生物群组成与人体测量和生活方式指标结合时,也被证明可以改善对特定餐后血糖反应的预测,其中Proteobacteria与较差的反应有关。个体对饮食的反应差异很大,而这种变化的决定因素一直难以捉摸。这些研究表明,可以针对个体的微生物群定制饮食,以最大化临床效果。
身体活动与体重管理
与饮食类似,身体活动是影响肠道微生物组的体重管理的关键因素,它通过多种途径影响肠道微生物组。虽然饮食可以通过改变肠腔营养环境直接塑造肠道微生物群,但身体活动对肠道微生物群的影响大多是间接的。相应地,身体活动对肠道微生物群组成的影响大小通常低于饮食(图5b)。
在啮齿动物中,肠道微生物群组成对急性和慢性身体活动都很敏感。然而,运动对肠道微生物特征的影响在研究中差异很大,这可能是由于运动类型和强度、饮食、物种和/或品系、年龄以及其他研究设计元素的差异。即使是自愿的轮跑与被迫的跑步机运动也会不同地改变小鼠的肠道微生物群。在迄今为止报告的效果中,相对可重复的是自愿轮跑对肠道微生物产生丁酸的能力的影响,包括在运动动物与久坐对照组中产生丁酸的类群的丰度更高和/或粪便或盲肠中丁酸浓度增加。
人类的肠道微生物群在活跃和久坐的人之间也有所不同。人类耐力跑者和更久坐的对照组在基线时肠道微生物群不同,在距离赛跑期间也会发生动态变化,特别关注的是Veillonella和Coriobacteriaceae类群的丰富。同样,精英划船运动员和超级马拉松运动员在运动前后的微生物群发生变化,橄榄球运动员与更久坐的个体相比表现出不同的肠道微生物分类和功能特征,职业和业余竞技自行车运动员之间的微生物群也有微小差异。几项研究报告了肠道微生物群结构和功能与心肺健康之间的相关性,通过氧气摄取量(V.O2max或V.O2peak)进行评估。此外,几项研究发现,与更久坐的个体相比,运动员的SCFA浓度和/或产生丁酸的类群有所增加,以及心肺健康程度较高的个体与较低的个体相比,这与动物模型的数据一致,表明运动丰富了肠道微生物群以产生SCFA。最近的一项研究报告称,产生SCFA的Bacteroides uniformis的绝对丰度与3000米赛跑时间相关,针对其丰度的饮食干预导致性能提高,这可以通过向小鼠施用B. uniformis来复制。同样,患有糖尿病前期的个体如果拥有更高基础SCFA产生能力的微生物群,在经过12周高强度运动干预后血糖反应的改善更大。适度活动似乎也以类似的方式影响肠道微生物群组成,活跃的女性拥有两种丁酸生产商Faecalibacterium prausnitzii和Roseburia hominis的水平增加,以及调节代谢的Akkermansia muciniphila(框2)。
然而,这些差异在多大程度上受到身体成分和其他生活方式因素如饮食的影响尚不清楚。的确,与更久坐的个体相比,精英橄榄球运动员的日常蛋白质摄入量增加解释了观察到的肠道微生物群组成差异的许多方面。一些纳入了严格饮食控制的研究报告了对身体活动的不同和可塑的肠道微生物反应。例如,在之前久坐的成年人中,在样本收集前实施了标准化的3天饮食干预,接受了6周的监督性耐力运动,干预终点的肠道微生物群的变化基于基线宿主表型而不同。运动增加了粪便丁酸和乙酸浓度以及产生丁酸的类群的相对丰度,这在瘦参与者中而不是在肥胖参与者中。同样,运动增加了瘦参与者中的Faecalibacterium spp.,但在肥胖参与者中则有相反的效果。在干预终点的微生物群变化在6周的久坐洗涤期后大部分被逆转,这表明肠道微生物群对运动输入的动态反应。
身体活动影响肠道微生物组的机制尚未阐明,但身体活动以多种方式改变了肠道腔环境。例如,运动改变了上皮内淋巴细胞中的细胞因子表达,这些细胞在调节宿主与肠道粘膜内微生物群之间的相互作用中起着关键作用。运动可能对肠道屏障完整性产生激素效应,剧烈运动会在短期内增加通透性,但常规运动促进降低通透性,正如训练有素的运动员与更久坐的个体相比,循环中的脂多糖水平较低。在高胆固醇血症小鼠中,自愿轮跑增加了主要胆汁酸分泌和粪便排泄,表明运动可能调节胆汁酸代谢。在无氧运动期间,循环中的乳酸可能转移到肠道腔,并改变腔道pH值。运动增加了活性氧的产生以及抗氧化酶活性,涉及活性氧的遗传和药理学操作的研究已经报告了对肠道微生物群多样性的影响。运动增加了肠道蠕动并减少了结肠传输时间,这与微生物群组成的改变有关。身体活动还短暂地提高了核心体温,引起了短期内对肠道血流的限制,改变了内分泌信号,产生机械力,并以预期影响肠道腔中竞争相互作用的方式改变了食物和水的消耗。
这些运动引起的肠道微生物组的变化是否改变了肠道微生物对能量代谢的贡献尚不清楚。一个挑战是宿主间反应的多样性,以至于即使研究检测到运动对肠道微生物组的影响,也可能报告没有队列范围内的代谢差异,可能因为应答者和无应答者效应。运动对肠道微生物组最明显的可重复效应,即更高的丁酸产生,可能会增加肠道屏障完整性,从而减少低度炎症,在过度营养的情况下带来代谢益处。因此,值得注意的是,接受来自控制饮食喂养的运动供体的粪便移植的高脂饮食喂养的肥胖小鼠表现出体重减轻,肝脏中促炎细胞因子表达降低,以及空腹血糖水平降低,同时增加了丁酸产生类群。然而,这些改善的代谢参数在多大程度上是由于运动、饮食或供体健康状况是不确定的,因为来自控制饮食喂养的运动供体的移植比来自高脂饮食喂养的运动供体的移植带来了更有益的结果,研究没有包括控制饮食喂养的非运动供体治疗,作者报告说饮食通常比运动对肠道微生物组有更强的影响。特别需要进行的研究是分离运动引起的肠道微生物组变化的能量后果,这将丰富我们对可用于微生物组定向体重管理的生活方式杠杆的知识。
临床路径与体重管理
当饮食、运动和其他行为干预措施被证明不足时,可能需要临床干预来管理体重及相关后遗症。在这一部分中,我们讨论了医疗干预与肠道微生物组之间的相互作用,以及针对肠道微生物组的体重管理临床疗法。
外科方法
治疗严重肥胖的外科方法旨在减少食物摄入和/或降低营养吸收。这些干预措施一直被证明可以增加微生物群的丰富度(α-多样性的衡量标准),以及增加变形杆菌门(Enterobacteriaceae)和Akkermansia属的相对丰度(补充表2)。这些与干预相关的微生物群组成变化在功能上与微生物代谢的变化以及通过证明与对照微生物群相比,无菌受体接受干预后肠道微生物群的减少脂肪量来展示与宿主健康的关联。不幸的是,有证据表明外科干预对微生物群的影响可能是短暂的,在干预后1年内有大量逆转。尽管如RYGB这样的外科方法总体上非常有效,但有一组患者在初始反应不佳或手术后体重重新增加。目前的证据表明肠道微生物群对RYGB后不同的反应有贡献是模棱两可的,研究一致认为肠道微生物群组成在经历成功和体重减轻效果不佳的个体之间只有轻微差异。然而,来自接受RYGB手术的人体粪便移植到经抗生素处理的小鼠表明,受体的体重增加表型与供体结果相关,说明肠道微生物群在一定程度上参与了减重成功。在经历体重减轻效果不佳的个体中观察到较高水平的Barnesiella属,以及它们的小鼠无菌受体,但需要进一步的实验来评估可重复性和因果关系。尽管研究处于早期阶段,现有的数据表明微生物群可以在一定程度上调节手术成功,提出了一个重要但未回答的问题:我们能否有一天仅通过精确修改肠道微生物群来模拟手术的效果?
药物
药物反应的个体间变异是医学中的主要挑战之一,可能部分由肠道微生物组介导。这可能通过多种机制发生,包括肠道微生物与口服药物的直接相互作用,常用药物对肠道微生物群的非靶向抗菌效应,以及药物对与肠道微生物群相互作用的宿主生理系统的影响。尽管这个新兴领域正在迅速扩展,它对体重管理和代谢健康的重要性是明确的。
也许,药物-微生物群相互作用在代谢健康中最著名的例子是二甲双胍,这是治疗2型糖尿病的一线疗法。值得注意的是,将经二甲双胍治疗的人体供体粪便移植到无菌小鼠中,与治疗前供体粪便相比,能够改善葡萄糖耐受性,提供了强有力的证据,表明肠道微生物群有助于药物效力。尽管二甲双胍耐受性良好并广泛使用,但约30%接受二甲双胍治疗的患者会出现与大肠埃希菌丰度相关的胃肠道副作用。或者,α-葡萄糖苷酶抑制剂阿卡波糖对肠道微生物群有相当大的非靶向效应。最近发现了一种高度普遍的微生物阿卡波糖抗性机制,导致药物失活,尽管其对药物效力的意义还有待于前瞻性研究中确定。
他汀类药物通常用于预防和控制心血管疾病,针对的是胆固醇生物合成中的HMG-CoA还原酶。在3个队列中的3000多名参与者中,他汀类药物的使用与炎症促进型微生物群特征呈负相关,其特征是高比例的拟杆菌属,低比例的Faecalibacterium属和低绝对微生物细胞密度。这些发现最近被复制并扩展,表明微生物群组成能够预测治疗反应,拟杆菌属富集、低多样性的微生物群不仅与增加的不良结果相关,而且与增加的治疗效应相关。即使在考虑到拥有更高基线肠道微生物群多样性的个体可能更健康和/或被开具了低效力他汀类药物剂量的可能性后,也发现具有更高基线肠道微生物群多样性的个体中他汀类药物的治疗效益较低,这表明更高的肠道微生物群多样性本身可能会妨碍药物效力。
粪便微生物群移植和其他恢复方法
许多有机体拥有调节代谢健康的冗余能力,这激发了通过粪便微生物群移植(FMT)转移整个微生物群落来实现治疗性操作的希望。FMT已被证明在对抗艰难梭菌的顽固感染方面有效,并在炎症性肠病方面显示出前景。然而,FMT在人类体重管理和肥胖方面的效果一直参差不齐,从没有临床效果到在结合饮食干预时对次要结果(如减少腹部脂肪和增加胰岛素反应)的积极影响。最近的一项研究检查了FMT是否可以在基于生活方式的干预后通过移植个体在最大减重点的微生物群来维持减重。肥胖或血脂异常患者通过运动指导和消费健康饮食、地中海饮食或绿色地中海饮食(地中海饮食加上绿茶和Wolffia globosa基奶昔)进行了为期6个月的生活方式减重。在接下来的8个月中,参与者被给予含有在6个月减重阶段末期收集的自体粪便的胶囊或安慰剂。只有在绿色地中海饮食组在减重阶段经历了肠道微生物群的显著变化,并且相应地,只有在绿色地中海组中,自体FMT对安慰剂相比减轻了体重重新增加、腰围增加和胰岛素反弹。这些数据表明,整个群落替代方法不太可能作为独立解决方案有效,特别是因为自体FMT绕过了异体FMT的一些更广泛的安全和生态系统匹配挑战,因此在很多方面代表了最佳情况。或者,微生物群恢复可以通过更有针对性的方式来实现,利用与健康紧密相关的明确定义的微生物群或微生物产品。尽管基于乳酸菌和双歧杆菌的传统益生菌和益生元对体重或葡萄糖稳态的影响似乎有限,新的候选物正在积极开发中。例如,在动物模型中施用巴氏杀菌的A. muciniphila似乎是安全的,并且可以通过对肠道屏障完整性的积极影响来防止饮食引起的肥胖、胰岛素抵抗和低度炎症。相应地,最近的一项40名参与者的探索性人类研究发现,每天口服10^10巴氏杀菌的A. muciniphila在3个月期间得到了良好耐受,显著降低了胰岛素抵抗和胰岛素血症,并与基线相比与身体脂肪和髋围的非显著减少相关(P = 0.09)。巴氏杀菌A. muciniphila的效力高于活细菌的发现提高了安全性,并确保了更长的货架稳定性。此外,最近的发现表明效力可能由外膜蛋白Amac_1100(参考文献133)或分泌蛋白P9(参考文献135)驱动,通过磷脂乙醇胺a15:0-i15:0(参考文献136)对稳态免疫的影响提供了额外的好处,为药物开发提供了靶点,并展望了更可预测的剂量动力学,为将基础发现转化为临床提供了一条路径。
走向微生物组感知的体重管理
在理解肠道微生物组对能量代谢影响方面的关键进展已经在动物模型中取得了,特别是小鼠,它们在解剖学、生理学、行为以及微生物组结构和功能方面与人类有几个不同之处。因此,实现肠道微生物组在改善代谢功能障碍方面的治疗前景将需要大量的转化研究。与动物模型相比,人类面临的强化挑战包括人类肠道微生物组中高度的个体间变异和纵向恢复力,低度的生态控制驱动了肠道微生物组在日常变化中的这些长期特征,以及发展可塑性在更长的人类寿命中施加影响,导致宿主表型和贡献微生物特征之间的时间脱节更大。开发协议和合成微生物群落,能够使动物宿主更忠实地复制人类-微生物组在能量代谢中的相互作用,将在缩小转化差距中至关重要。补充小鼠研究的还有在更大动物中的无菌研究,如猪,这些动物在生活史、生理学和微生物组方面更接近人类。体内方法,特别是涉及设计为模拟人类肠道生理条件的生物反应器,以及体外方法,如来自诱导多能干细胞或活组织检查的类器官,是重要的补充工具,具有更大的控制力和可重复性的优势。
在不同个体和环境中稳定操纵人类微生物组的困难表明,像FMT这样的一刀切和未经提炼的粗暴方法不太可能取得持久的成功。相反,通过针对特定肠道微生物功能和代谢产物的个性化干预,可能会取得更大的长期影响。在这样做的过程中,我们需要牢记细菌产品的多效性。例如,短链脂肪酸在结肠能量回收中的参与,同时也最小化饥饿,增加产热和调节胚胎组织发育(图2-4),提出了基于短链脂肪酸的干预可能具有复杂甚至代际的结果。面对这些挑战将使肠道微生物组的合理操纵能够持续足够长的时间来改善健康,并且要么在个体之间普遍适用,要么以公平的方式个性化。
我们预计在未来10年内,我们将揭示许多我们可以使用的微生物组调节杠杆。菌株水平鉴定、培养、基因组编辑和代谢物鉴定方面的持续技术进步将使我们能够更具体地描述感兴趣的微生物参与者和鼓励它们存在的生态条件。例如,尽管它们的16S rRNA基因序列是同质的,但最近的研究在A. muciniphila菌株中发现了大量全基因组差异,这可能有助于解释为什么菌株对炎症有不同的影响。继续阐明微生物组操纵能量平衡的机制预计将突出显示调节饥饿和饱腹感、饮食能量收获、能量分配以及能量代谢与炎症之间相互作用的有希望的干预目标。例如,利用次级胆汁酸通过TGR5介导的棕色脂肪组织激活的产热效应(图3和4),最近发现,通过补充非12α-羟基化胆汁酸或Parabacteroides distasonis(一种能够产生这些酸的细菌)可以抑制饮食限制后的体重反弹。此外,对细菌化学的快速进展知识,包括肠道微生物组与治疗药物之间的普遍相互,可能建议新的微生物组靶向方法,通过增强或抑制细菌生物转化来增强现有药物的效力。细菌异源代谢的新兴知识最终可能会影响有关药物选择、剂量和/或联合给药代理的临床决策,正如已经为地高辛、伊立替康和左旋多巴这样的非代谢药物提出的。相比之下,我们对小肠和粘膜相关微生物组的知识进展较慢,因为获取这些样本需要更大的侵入性。尽管如此,在小肠中,宿主和微生物直接争夺营养物质,在粘液层,宿主组织和微生物最为接近,这些相互作用对能量代谢和炎症最有影响。更多地关注这些微生物群落可能会揭示额外的、高影响力的调节目标。
迄今为止的大多数研究集中在超重和肥胖上,而不是营养不良。然而,现有的证据表明,肠道微生物组的破坏先于儿童营养不良的发生,与消瘦和发育迟缓的严重程度相关,并通过移植将体重损失表型传递给无菌小鼠。这些数据表明肠道微生物组在营养不良中起着关键作用,为开发以微生物组为重点的疗法提供了成熟的机会,以改善全球三分之一患有发育迟缓和/或消瘦的儿童的健康状况。尽管还有许多有待发现,但目前正在进行的有希望的微生物组重点临床试验针对与营养不良或超重相关的代谢表型。例如,研究人员最近开发了几种微生物群定向辅助食品(MDCF)配方,这些配方被评估了它们成熟营养不良的孟加拉国儿童的肠道微生物组到健康断奶后概况的能力。领先的MDCF原型随后在孟加拉国幼儿中显示出比标准即食治疗食品更好的功效,这些幼儿患有中度营养不良。
尽管还有许多有待发现,但目前针对与营养不良或超营养相关的代谢表型的有希望的微生物组重点临床试验正在进行中。例如,研究人员最近开发了几种微生物群定向辅助食品(MDCF)配方,这些配方被评估了它们成熟营养不良的孟加拉国儿童的肠道微生物组到一个健康的断奶后概况的能力。领先的MDCF原型随后在孟加拉国幼儿中显示出比标准即食治疗食品更好的效果,这些幼儿患有中度急性营养不良,引发了体重-身长和体重-年龄z得分的显著相对改善,以及影响骨骼生长和神经发育的血浆蛋白的变化。同样,如前所述,最近的一项研究发现,饮食是自体粪便微生物群移植(FMT)在减轻肥胖或血脂异常患者体重重新增加方面的效力的重要支持,这些患者经历了6个月的生活方式驱动的减重,因为FMT只有在与特定的绿色地中海饮食结合时才有益。
与MDCF一样,这些数据表明饮食可能作为微生物组靶向疗法的辅助手段。未来的微生物组靶向疗法的人体试验应该通过提供含有支持微生物植入和有益微生物生长的基质的固定组成来标准化参与者的饮食。对饮食加工的更多关注也有助于标准化向结肠中最密集的微生物群落提供营养素。尽管如此,独立的饮食操作途径也显示出前景,例如一项研究报告称,巴氏杀菌的A. muciniphila的益生菌管理或传递特定的Akkermansia衍生效应分子,如Amac_1100,可以减少脂肪和胰岛素抵抗。总的来说,这样的试验作为重要的概念验证,表明微生物组意识疗法可以与现有的体重管理方案相比,并对其进行补充。
往期推荐
