

论文ID
本文译自:Schupack DA, Mars RAT, Voelker DH, Abeykoon JP, Kashyap PC. The promise of the gut microbiome as part of individualized treatment strategies. Nat Rev Gastroenterol Hepatol. 2022 Jan;19(1):7-25.
发表杂志:Nat Rev Gastroenterol Hepatol
影响因子:67.5
通讯作者:Purna C. Kashyap
作者单位:Mayo Clinic, Rochester, MN, USA.
摘 要
疾病表现、进展和治疗反应的变异性一直是医学领域的一个中心挑战。尽管宿主因素和遗传学的变异性很重要,但现在已经变得明显的是,在迈向个体化治疗的过程中,必须考虑具有巨大遗传和代谢多样性的肠道微生物群。在这篇综述中,我们讨论了六大广泛的疾病组:传染病、癌症、代谢性疾病、心血管疾病、自身免疫或炎症性疾病以及过敏性和特应性疾病。我们强调了当前对肠道微生物群在疾病发病机制和预后、疗效、治疗相关不良事件中的知识,并强调了它在分层现有治疗和作为新疗法来源方面的前景。这篇综述不打算对每种疾病状态进行全面介绍,而是突出了微生物群作为临床实践中个体化治疗策略工具的潜在意义。尽管还为时尚早,但前景是乐观的,但在临床实施之前需要克服挑战,包括提高对潜在机制的理解、进行反映肠道微生物群和宿主反应的多层数据的纵向研究、标准化的测试和报告方法,以及在更大的队列中进行验证。鉴于微生物群领域与基础和临床研究的同时进展,微生物群很可能在未来十年内成为临床治疗的一个组成部分。
正文
个体间在疾病表现、治疗反应和治疗不良效应方面的变异性,为疾病有效管理和患者安全带来了重大挑战。这种认识是精准医学的基础,其最简单的形式可以被描述为使用个体化方法,不通过试错就为正确的患者确定正确的治疗方法。精准医学领域的初始范例集中在基于宿主遗传学和一系列临床及实验室特征对患者进行亚型分类,例如,肿瘤组织的遗传构成。这一过程在预测疗法的疗效和毒性方面取得了显著改进。然而,宿主基因组仅代表超有机体基因的一部分,因为我们体内微生物群落的遗传和化学多样性大于人类基因组中编码的多样性。由于个体拥有独特且广泛变化的肠道微生物群,它对疾病表型的异质性以及治疗反应的贡献也就不足为奇了。
肠道微生物群与人类遗传学的一个不同之处在于,它代表了我们健康的一个动态组成部分,通过复杂的网络不断与宿主和环境因素相互作用。尽管这可能构成一个挑战,但肠道微生物群的可塑性也提供了一个独特的机会,使其成为精准医学的一个有吸引力的目标。目前存在几种类别和水平的基于微生物群的治疗,针对微生物群的特异性各不相同。粪便微生物群移植(FMT)是涉及将一个人的粪便整体转移到另一个人身上,以尝试用“健康”的微生物群重新殖民肠道。益生菌(在适当管理下假定对健康有益的活微生物)有针对性治疗的潜力,尽管临床实践中使用的大多数当前配方缺乏所需的特异性。益生元是被选择性消化以刺激有益生物生长或活动的化合物,因此对治疗的个性化有很大的潜力。合生元是微生物和底物的混合物,它们被宿主选择性利用以实现某些健康益处,为微生物群相关治疗的个性化提供了更大的空间。除了以纯化形式补充特定化合物外,饮食摄入富含益生元化合物的食物是改变微生物群种群的另一种可能途径。关于特定微生物群驱动的机制在疾病发病机制、预后和治疗结果方面的累积数据,已经导致调查了多种额外的方法,包括但不限于工程细菌以执行特定功能、改变微生物功能或其与宿主相互作用的小分子药物和生物制剂,以及改变微生物组成和功能的原生或工程化的噬菌体。
在这篇综述中,我们总结了当前支持使用肠道微生物群作为精准医学工具的证据,并建议未来需要进行的工作,以将微生物群作为一种工具纳入个体化治疗或干预措施。我们选择了六个有相对强有力的证据支持肠道微生物群作用的广泛疾病组。尽管在每个疾病组中都有激动人心的发展,但当考虑到临床影响时,不同疾病组的前景和成熟度水平各不相同(见图1;补充图1)。我们从临床实践知识成熟度最高到最低的疾病状态进行讨论。在每个部分中,我们旨在提供有关该疾病状态下已知的分类学关联和微生物群机制、当前使用的微生物群导向治疗、肠道微生物群在预测治疗反应和毒性中的作用,以及对新型微生物群治疗的展望的信息。鉴于当前这些疾病状态关于肠道微生物群的证据存在差异,这种组织方式并不是完全统一的。我们的目标不是提供每个疾病状态下肠道微生物群的详尽概述,而是突出与精准治疗领域相关的研究,并综合当前状态和这一快速发展领域的成熟度。综述提供了令人信服的证据,支持将微生物群纳入临床实践,同时承认挑战并概述了实现这一目标的途径。
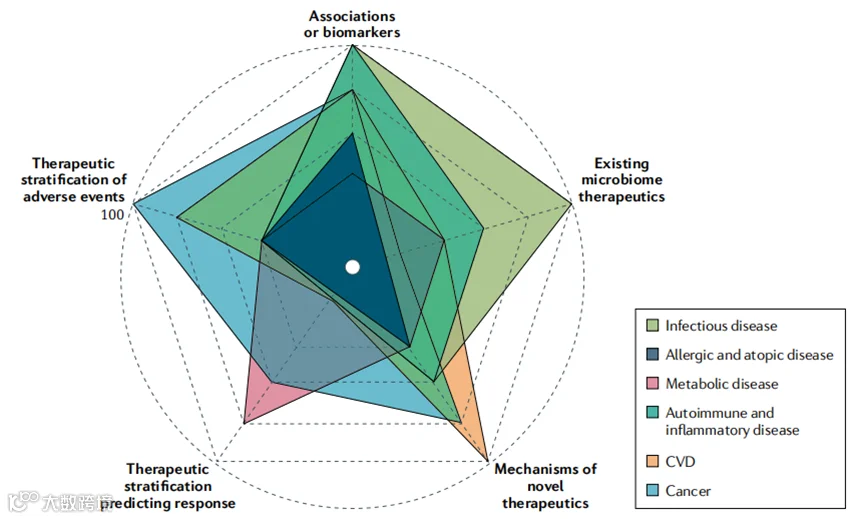
图1 | 肠道微生物群作为精准医学工具的成熟度。尽管微生物群科学的进展总体上令人瞩目,但在不同疾病状态中的进展并不均衡。在本图中,我们根据对人类可用文献的综述,概述了微生物群领域在各个疾病组向临床实践进展的成熟度。我们在每个疾病组内考虑以下参数:存在稳健的分类学关联或生物标志物;微生物群治疗的存在;可操作的微生物群机制的知识;微生物群在治疗分层中的使用;微生物群在预测不良治疗效果中的使用。CVD,心血管疾病。
传染病
抗生素引起的肠道微生物群破坏,促进机会性感染和医院获得性感染的机制,是一个引起强烈兴趣的话题。我们以艰难梭状芽胞杆菌(Clostridioides difficile)为例,这是最常见的医院获得性腹泻感染,来突出微生物群和病原体特有的特征,这些特征可能解释了临床结果中的个体间变异性。复发性艰难梭状芽胞杆菌感染(CDI)是微生物群研究的一个中心焦点。CDI最常发生在抗生素使用后,而矛盾的是,CDI的一线治疗也包括抗生素。尽管抗生素在人群层面上相当有效,但部分患者要么治疗失败,要么在成功治疗后CDI可能会复发,这可能与宿主特征有关,如高龄,或与使用质子泵抑制剂等药物,以及在肠道微生物群的背景下特定病原体的特征有关。
除了宿主因素,肠道菌群的破坏也被发现是CDI(艰难梭状芽胞杆菌感染)背后的一个关键因素。研究发现,与健康个体作为对照相比,CDI患者体内肠球菌(Enterococcus)、韦荣氏球菌(Veillonella)、乳酸杆菌(Lactobacillus)和伽马变形菌(Gamma-proteobacteria)种类的相对丰度升高,而拟杆菌(Bacteroides)、毛螺菌科(Lachnospiraceae)和瘤胃菌科(Ruminococcaceae)种类的水平较低。此外,在无菌小鼠中进行的模型研究显示,由一系列宿主因素驱动的肠道微生物群的改变增加了CDI的易感性,原因是氨基酸的可用性增加,这是艰难梭状芽胞杆菌偏好的营养生态位。同样,其他使用小鼠模型的研究发现,微生物群衍生的抑制因子的丧失以及CDI中开放营养生态位的增加,这包括短链脂肪酸(SCFAs)(特别是戊酸)和次级胆汁酸脱氧胆酸的水平下降,以及有机酸琥珀酸、唾液酸和氨基酸水平的增加。这些增加CDI易感性的微生物群驱动因素在个体之间是不同的,并非每个发展为CDI的个体都表现出所有这些异常。
除了肠道微生物群,艰难梭状芽胞杆菌(C. difficile)菌株之间的变异,如毒素产生、代谢和生物膜形成能力的变化,可能会导致不同的结果。一项使用全基因组测序研究了约400个来自CDI(艰难梭状芽胞杆菌感染)患者的临床分离株,发现大多数疾病复发是由与初次感染相同的菌株引起的,这表明使菌株能够在肠道中持续存在的特性可能与复发性疾病有关。艰难梭状芽胞杆菌组织成多细胞生物膜可能允许持续存在,因为生物膜可以提供对抗生素的物理屏障,并能干扰从胃肠道的清除。因此,结合全基因组测序、肠道中的代谢环境和特定微生物类群的诊断测试,在考虑个体化治疗方法时可能会提供更高的分辨率。
目前治疗多次复发性CDI(艰难梭状芽胞杆菌感染)或治疗失败的方法是粪便微生物群移植(FMT),它在治疗复发性CDI方面非常有效,无论采用何种给药途径、植入程度或供体肠道微生物群的特定特征。预测FMT失败的主要特征是持续使用抗生素。尽管FMT具有疗效,但人们担心其长期安全性,目前正在一个长期登记处进行调查。此外,关于如何最好地筛选供体粪便的问题仍然存在。应该使用宏基因组学或病原体全基因组测序来识别抗生素耐药性档案,还是使用培养或基于PCR的病原体筛选就足够了?全基因组测序方法的有用性通过两项临床试验得到说明,这些试验涉及与供体粪便相关的产生广谱β-内酰胺酶的大肠埃希菌引起的菌血症病例。FMT的这种安全性问题对于免疫受损的患者尤其相关。
除了粪便微生物群移植(FMT),肠道微生物群也被发现可以预测CDI(艰难梭状芽胞杆菌感染)的治疗结果。在分类学层面,治疗前瘤胃菌科(Ruminococcaceae)、拟杆菌科(Rikenellaceae)、拟杆菌属(Bacteroides)和费卡利杆菌属(Faecalibacterium)的丰度与CDI对抗生素的积极反应相关,而梭菌科(Clostridiaceae)、毛螺菌科(Lachnospiraceae)、布氏菌属(Blautia)、粪球菌属(Coprococcus)、链球菌属(Streptococcus)、双歧杆菌属(Bifidobacterium)、瘤胃菌属(Ruminococcus)和放线菌属(Actinomyces)的更高水平与治疗无反应相关,这是一项包括88名CDI患者的研究。其中一些类群还预测了复发感染的风险,而其他类群则与艰难梭状芽胞杆菌无法在肠道中定植的总体能力相关(见图2)。
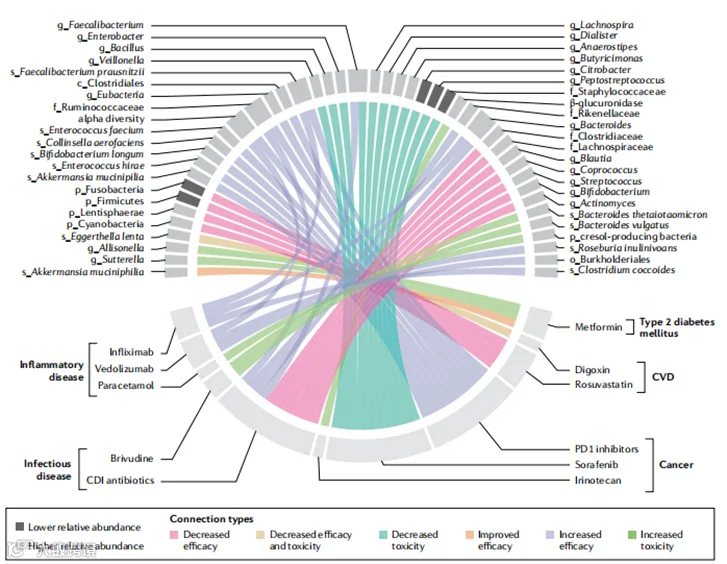
图2 | 肠道微生物群与药物治疗之间的关联。特定微生物类群与常用药物的疗效或毒性之间的关联。本图仅包括人类研究,我们不考虑基于单独关联或额外生物学验证的发现中置信度水平的差异。前缀表示系统发育水平(p_门,o_目,f_科,g_属,s_种)。还有许多其他药物,其体外或动物数据表明与结果有关联,我们预计这一领域将迅速扩展。CDI,艰难梭状芽胞杆菌感染;CVD,心血管疾病。
对微生物群驱动的CDI潜在机制的深入理解,以及肠道微生物群在治疗结果中的预后作用,结合对FMT长期风险的不确定性,已经促进了针对CDI的基于微生物群的诊断和治疗的激增。其中最先进的是定义的微生物群落,在II期临床试验中显示出前景,并且现在已在III期临床试验中取得成功。此外,还有几种利用特定细菌和/或代谢产物、益生元和噬菌体的方法正在早期研究阶段。这些范围较窄的治疗方式将可能允许基于宿主和宿主微生物群特征的更个性化的治疗,可能会进一步提高治疗CDI的疗效并降低风险。这一进展为开发未来的、更精确的基于微生物群的治疗策略在CDI中提供了乐观的前景(见表1)。
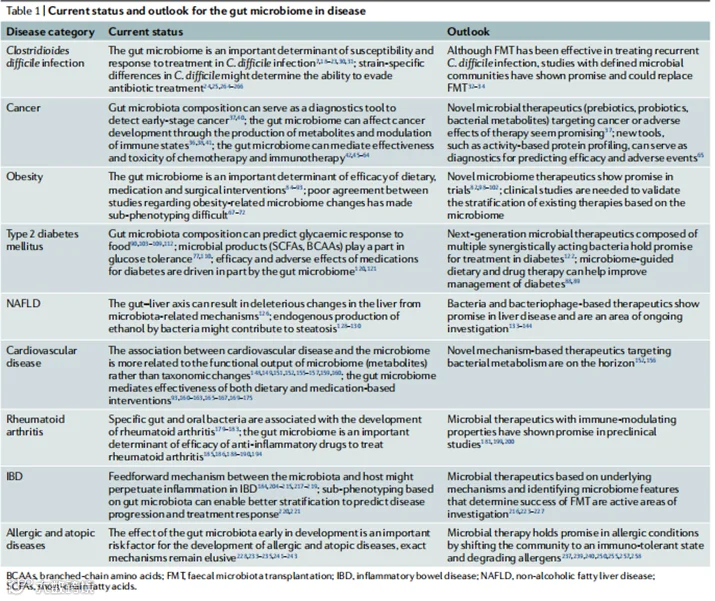
表1 肠道微生物组在疾病中的研究现状与展望
肿瘤
肠道微生物群可以从四个不同的视角为个性化癌症生物学领域提供信息:抑制癌症发展、识别新的治疗方法、优化现有治疗以及癌症诊断。在癌症发展和识别新治疗方法方面,这些问题与肠道微生物群-免疫系统轴的不完全功能有关,肠道微生物群和肿瘤组织本身对免疫系统既有免疫抑制作用也有免疫刺激作用。优化现有治疗与减少不良效应和预测对癌症治疗的反应有关,这些治疗方法包括化疗、放疗到新的免疫治疗方法。在这里,肠道微生物群在药物代谢以及影响免疫细胞和细胞因子水平方面的作用,可能导致治疗反应的变化以及不良效应的发展。
微生物群与癌症的联系
肠道微生物群-免疫系统轴的机制知识可以被探索,以拦截癌症发展并识别新的治疗方法。癌症发展和疾病进展可以通过微生物的致癌效应及其产物来影响,调节可能促进或抑制肿瘤生长的循环代谢物水平,并诱导促炎和免疫抑制效应。与涉及器官(如结直肠、宫颈、肺、头颈)的癌症发展和进展程度相关的微生物群的局部改变已被识别。肠道微生物群与癌症之间的机制联系在结直肠癌中最为明显。微生物及其代谢产物与肠粘膜的复杂关系可以导致粘膜通透性的变化,增加局部暴露于多种潜在致癌化合物的机会,并可能导致慢性炎症状态。另一个机制联系的例子是携带致病性岛pks的大肠杆菌菌株,该岛编码合成基因毒性次级代谢产物colibactin的基因,可以用来预测结肠癌风险。除了明确的机制特征外,肠道微生物组成和功能属性的差异已与器官的恶性程度相关,这些器官与肠道微生物群的直接接触较少。例如,涉及68名胰腺癌患者的研究表明,肿瘤组织中假黄单胞菌属(Pseudoxanthomonas)、糖多孢菌属(Saccharopolyspora)和链霉菌属(Streptomyces)的丰度与手术切除后的长期生存相关,与那些预后较差的患者相比。这些保护效应通过小鼠模型的粪便移植被转移,表明粪便微生物群移植(FMT)作为一种潜在的治疗方式需要更广泛的考虑,至少作为辅助形式。
微生物群与癌症治疗
肠道微生物群可以影响化疗和免疫疗法的反应,无论是在疗效还是毒性方面。这种关系可能是由于在抗原呈递、诱导炎症反应以及药物化学修饰方面的协同效应。例如,环磷酰胺是一种广泛使用的烷化剂,它通过诱导DNA交叉链接,部分通过调节与特定T辅助细胞17(TH17)反应生成相关的免疫途径来施加其抗肿瘤效应。在小鼠模型中,细菌转移到次级淋巴器官后,与此相关的免疫途径会生成特定的T辅助细胞17(TH17)反应。在一项关于诱导类型1免疫反应的CpG寡脱氧核苷酸和DNA交叉链接剂奥沙利铂的小鼠研究中,抗生素存在的情况下治疗效果降低,这归因于肿瘤相关骨髓细胞产生的肿瘤坏死因子(TNF)减少。鉴于奥沙利铂的治疗效果部分是由肠道微生物群产生的活性氧物种驱动的,因此抗生素引起的奥沙利铂疗效降低可能是多因素的。最后,通过细菌胞嘧啶脱氨酶的作用,对吉西他滨进行微生物代谢,生成2',2'-二氟脱氧尿苷,可能会降低其治疗效果。
除了化疗药物外,越来越多的证据表明肠道微生物群在决定癌症免疫疗法的有效性方面的重要性。在人类中,使用抗生素与免疫疗法治疗多种癌症类型(非小细胞肺癌、肾细胞癌和尿路上皮癌)时,与减少无进展生存期和总生存期相关。这一发现通过动物模型进一步调查,表明抗生素这些负面影响中免疫介导机制的作用。在小鼠中,当拟杆菌属的B. fragilis和B. thetaiotaomicron以及Burkholderiales的丰度增加时,CTLA4抑制剂ipilimumab的疗效得到改善,这与IL-12依赖的TH1免疫反应的上调有关。在小鼠结肠癌模型中,当Bifidobacterium pseudolongum存在时,通过免疫激活增强了CTLA4阻断的疗效,这是由于治疗导致的粘膜屏障功能降低,从而促进了肌苷的增加转移。IL-12还被发现在小鼠上皮肿瘤中CCR9+CXCR3+CD4+细胞的招募中发挥作用,这反过来与人类肠道微生物群中Akkermansia muciniphila的增加丰度相关。在小鼠中,随着Bifidobacterium种群的增加,PD1和PDL1检查点抑制剂对黑色素瘤的疗效也可以得到增强。有趣的是,即使在没有常规治疗的情况下,双歧杆菌属也可能具有抗黑色素瘤的效应,因为在同一群小鼠中,治疗前丰度与肿瘤生长抑制相关。这种效应被认为与树突状细胞功能的上调有关,导致肿瘤微环境中CD8+ T细胞的活性和积累增强。在42名转移性疾病患者的研究中,PD1抑制剂治疗黑色素瘤的疗效也在Bifidobacterium longum、Collinsella aerofaciens和Enterococcus faecium的丰度增加的情况下得到增强。而在另一组43名黑色素瘤患者的人类队列中,微生物群多样性的增加与PDL1和PD1治疗后癌症生存率的提高相关。具体而言,与无反应者相比,反应者的病人中瘤胃菌科、Firmicutes、Eubacterium sp.、Clostridia、Clostridiales和费卡利杆菌属(Faecalibacterium prausnitzii)的丰度增加(见图2)。
化疗药物的毒性可能是决定其使用能力的主要因素。肠道微生物群可以通过几种方式改变药物代谢,包括竞争性抑制、肠道微生物的直接代谢效应以及改变宿主基因表达,这些基因涉及代谢途径,如在无菌小鼠中观察到的外源性解毒基因的下调。具体来说,抗癌药物如5-氟尿嘧啶(5-FU)、伊立替康和索拉非尼的毒性被认为与肠道微生物代谢有关。当与病毒DNA聚合酶抑制剂索立夫定在大鼠模型中共用时,DNA复制器5-FU的毒性是由于索立夫定转化的微生物产物溴乙烯尿嘧啶(BVU)引起的5-FU代谢减少。了解负责BVU形成的特定生化途径的知识,可以帮助预测这种毒性,以通知替代治疗选择或开发特定的BVU形成抑制剂。拓扑异构酶抑制剂伊立替康的使用通常受到严重腹泻的发展阻碍,这与细菌β-葡萄糖醛酸酶介导的非活性伊立替康代谢物的重新激活有关;抗生素治疗已被证明可以减少活性伊立替康代谢物的体外产生,给予降低β-葡萄糖醛酸酶活性的益生菌混合物可以改善结肠癌患者的腹泻,并且β-葡萄糖醛酸酶的小分子抑制剂在临床前小鼠模型中显示出前景。酪氨酸激酶抑制剂索拉非尼是另一种毒性可能与肠道微生物活动有关的药剂,因为在肝细胞癌患者中使用索拉非尼后出现的腹泻和手足皮肤反应与特定的微生物类群有关。具体来说,Veillonella、Bacillus、Enterobacter、Faecalibacterium、Lachnospira、Dialister和Anaerostipes的丰度对手足皮肤反应有保护作用,Butyricimonas的丰度和Citrobacter、Peptostreptococcus和Staphylococcaceae的较低水平与腹泻的发展较少有关。这种效应的潜在机制可能通过药物及其代谢产物的肠肝循环而减弱。一项对接受其他酪氨酸激酶抑制剂(帕唑帕尼和舒尼替尼)治疗的转移性肾细胞癌患者的研究表明,与安慰剂相比,使用来自健康供体的FMT治疗可以改善治疗引起的腹泻,进一步涉及微生物群改变与这种不良反应的关系。开发基于活性的蛋白质探针以识别负责外源性物质代谢的特定微生物途径,作为诊断工具具有前景,并可能实现更好的治疗分层。
新型治疗可能使用细菌菌株或纯化的病原体相关分子模式作为Toll样受体(TLR)激动剂,以触发TLR刺激水平较低的患者的局部免疫反应。此外,可以影响肿瘤生长的粪便和循环微生物代谢物水平(短链脂肪酸、次级胆汁酸、维生素和多胺)可以用来评估治疗前的代谢健康状况,进而影响下一代生物治疗药物,可能与饮食干预相结合。一系列免疫细胞亚群(TH17细胞、调节性T细胞1型、细胞毒性T淋巴细胞、CD4+细胞、CD8+细胞)和细胞因子丰度(通过芳香烃受体(AHR)信号传导的TNF、IL-12、IL-22)受到肠道微生物群变化的影响,可能被调节以影响癌症免疫监视。这些标记的测量还可以用于评估患者的免疫健康状况,并提供干预目标。这些方法将需要在大型队列上进行详细的个性化多组学研究,然后才能在临床中得到应用。
迄今为止的研究为肠道微生物群在癌症表型的异质性和对癌症治疗的反应中的作用提供了有力支持。然而,来自人类研究的数据主要是关联性的,仍然需要在不同的队列中进行复制。机制数据主要来自动物研究,这引发了关于其转化有效性的一些担忧。尽管存在这些担忧,但前景依然乐观,将微生物群作为癌症治疗策略的一个组成部分进行整合似乎是不可避免的(见表1)。
代谢性疾病
肥胖
超重或肥胖的儿童和成人数量正在增长,美国超过40%的成年人现在符合肥胖标准。应对肥胖的一个主要挑战是机制的复杂性,这是由遗传、肠道微生物群、饮食和环境之间的相互作用导致的,这些相互作用导致生理变化,促成了肥胖。
微生物群与肥胖的联系。2016年发表的一篇对精选的人类微生物群研究的元分析发现,肥胖与样本内alpha多样性指标的丰富度和均匀度的降低有小但统计学上显著的关联。除了alpha多样性,早期的微生物群研究在小鼠模型和人类研究中都报告了肥胖中拟杆菌门(Firmicutes)与杆菌门(Bacteroidetes)比例升高的情况,但在这项元分析中并未复制出来。尽管缺乏稳健的组成标记,肠道微生物群在肥胖中的作用得到了粪便移植实验的支持,这些实验表明,由肥胖不一致的单卵双胞胎的粪便定植的无菌小鼠表现出供体的代谢表型。这一前提因以下观察而得到进一步加强:与低脂植物多糖饮食相比,由高脂饮食引起的体重增加在人源化小鼠(用健康人类粪便定植的无菌小鼠)中可以通过仅仅移植这些人源化小鼠的粪便样本传递给接受者无菌小鼠,而不需要持续喂食高脂饮食。这一发现表明,即使肥胖表型是由其他原因引起的,微生物群在传播肥胖表型中也很重要。肠道微生物群引起的肥胖的几种机制包括从饮食中提高能量提取效率、影响饱腹感和能量摄入、全身炎症和胰岛素抵抗。
在肥胖中缺乏一致的组成标记表明存在大量的功能冗余。实际上,在功能层面上冗余较少,因为几种不同的微生物群落配置的转变可以驱动一系列生物活性因子的产生变化,如短链脂肪酸(SCFAs)、胆汁酸和脂多糖(LPS),所有这些都与肥胖有关。SCFAs在激素信号传递中发挥作用,例如血清素和肽YY的释放,这在饱腹感中发挥作用,暗示了肠-脑轴在推动肥胖中的参与。尽管在理解微生物群驱动的肥胖潜在机制方面取得了重大进展,但我们尚未有足够的分辨率来根据潜在的微生物群基础机制对肥胖个体进行分层。随着这一领域证据的积累,人们可以轻易设想,基于微生物群的个体分层将成为未来个性化肥胖管理策略的一部分。益生菌作为肥胖治疗方案的有效性已在动物模型中被提出,但人类临床试验的结果混杂,鉴于所使用的益生菌配方缺乏一致性,其作用在这一点上仍然不清楚。关于使用益生元进行治疗,同样在动物模型中已有令人鼓舞的发现,但就持久减重而言,尚未在人类临床试验中明确转化为人类应用。
微生物群与肥胖治疗
肥胖管理的主要治疗方法是饮食、药物(如胰高血糖素样肽1(GLP1)激动剂、奥利司他和苯丙胺)和减肥手术。饮食可以在短期和长期内影响肠道微生物群,对短暂的饮食变化作出反应时会发生改变。更重要的是,饮食的治疗效应也依赖于个体的微生物群。在将肠道微生物群简化为无菌小鼠中的10种微生物菌株时,已经证明饮食改变可以改变肠道的定植模式及其发酵能力。在肥胖个体中进行饮食改变的一种方法是增加水果、蔬菜和低能量密度食品的摄入量,同时减少高营养密度食品的摄入量;然而,对这种干预的反应非常多变。一项针对26名超重或肥胖个体的初步研究发现,在进行这种饮食干预之前,肠道微生物群中预测的糖苷水解酶高丰度与体积饮食干预后体重减轻<5%相关,这表明基线肠道微生物群在预测结果中可能发挥作用。这些发现与个体间餐后血糖反应和血脂水平的变异性一致,这已被归因于肠道微生物群。在21名健康个体(包括正常BMI(>18.5和<25)和高BMI(≥30)的个体)的人群中,增加的卡路里摄入量与肠道微生物群的快速改变相关,在3天内,包括个体与拟杆菌门(Firmicutes)的相对丰度增加和杆菌门(Bacteroidetes)的耗竭。这些变化与粪便热量含量的减少有关,表明增加能量收获。因此,减少卡路里摄入量可能通过改变肥胖个体亚群的微生物群而产生有益效果。肠道细菌的相对丰度,如Eubacterium ruminantium和Clostridium felsineum,也与78名肥胖个体对几种可变饮食干预的微生物群可塑性增加相关。总的来说,这些研究表明,一些食物对每个人都是“健康”的概念过于简单,基于肠道微生物群指标的饮食选择可能在体重管理方面有益。
很少有研究探讨了肠道微生物群在针对肥胖的药理学和手术治疗背景下的作用。然而,发现GLP1激动剂利拉鲁肽(它在血糖水平升高的情况下增加胰岛素释放并延迟胃排空)在大鼠中增加了拟杆菌门(Firmicutes)与杆菌门(Bacteroidetes)的比例,因此可能至少部分通过肠道微生物群的次级变化来促进减重。之前进行的Roux-en-Y胃旁路手术(一种用于管理那些未能通过生活方式改变和药物治疗的患者)与术后15个月拟杆菌门相对丰度的减少和如变形菌门(Proteobacteria)等兼性厌氧菌水平的升高相关。这些变化可能在减重中发挥作用。另一种手术——袖状胃切除术——在减少炎症和改变肠道微生物群方面是有效的,它使23名术前肥胖患者的肠道微生物群更接近作为对照的健康个体,伴随着由微生物群决定的血浆谷氨酸水平的恢复,作为肥胖的生物标志物。虽然大多数发现需要在更大的队列中验证并在机制模型中测试,但这些研究突出了微生物群分析在评估当前可用的肥胖疗法的有效性方面的实用性。
在基于微生物群的治疗方面,阿克曼氏菌(A. muciniphila)是治疗代谢综合征和肥胖的有希望的候选者。在小鼠中,由于改善了肠道屏障功能、减少了内毒素血症并通过激活TLR信号传导改善了葡萄糖耐量(由于外膜蛋白),食用阿克曼氏菌对体重增加具有保护作用。在32人的初步数据中显示了阿克曼氏菌的安全性和有效性,在3个月内实现了适度的体重减轻和肥胖实验室标记的改善。同样,克里斯滕森氏菌(Christensenella minuta),被认为是最具遗传性的微生物种类之一,并且特别与人类的瘦体型相关,在动物模型中也被发现对治疗肥胖有效,并且计划很快开始在人类中进行随机对照试验。尽管我们强调了一些针对微生物群以减少肠道衍生的炎症信号的正在进行的努力,但改变营养信号和调节肠-脑轴也可能是有效的策略。这些针对微生物-宿主相互作用的新方法很可能是预防和治疗肥胖的重要组成部分(见表1)。
胰岛素抵抗或糖尿病
由于对循环代谢物和免疫状态的影响,胃肠道及其相关的肠道微生物群可以类似地被视为一个内分泌器官。因此,它通过影响胰岛素信号传导参与葡萄糖代谢。
微生物群与改变的血糖控制的联系。几项研究提供了糖尿病中肠道微生物群的横向分类学变化,表现为丁酸产生菌的丰度降低和整体微生物多样性的减少,但研究发现在各项研究中并不一致。然而,已经阐明了肠道微生物群与胰岛素抵抗之间的明确关系,来自代谢综合征供体的粪便微生物群移植(FMT)与来自Roux-en-Y胃旁路手术供体的FMT相比,会导致无菌小鼠的胰岛素敏感性降低。另一项人类研究进一步支持了这一点,在进行干预之前,来自瘦供体的FMT改善了肥胖个体的胰岛素敏感性。这突出了针对肠道菌群的治疗在经过仔细选择的患者中的潜在治疗益处,对于这些患者,肠道微生物的转变可能会导致更实质性的临床益处。在人类的随机临床试验中显示,增加多样性和丰度的一组由膳食纤维促进的SCFA(短链脂肪酸)产生菌导致了血红蛋白A1c水平的改善,这归因于胰高血糖素样肽的增加。然而,其他潜在的SCFA产生者减少或保持不变,这表明并非所有的SCFA产生者都是平等的,更有针对性的恢复特定微生物可能更有益。未来的研究将有助于识别特定的细菌群,它们不仅能够执行功能,实际上还能协同工作以恢复关键功能。总的来说,这些研究支持了肠道微生物群与糖尿病之间的联系,并突出了利用肠道微生物群优化治疗的前景。肠道微生物群在胰岛素抵抗或2型糖尿病(T2DM)中的作用机制与肥胖所确定的机制重叠,例如低度炎症、胃肠通透性的变化可能伴随内毒素血症、SCFA产生和吸收的减少,这与代谢综合征的概念一致。SCFA的变化反过来又可以影响各种代谢激素的产生,如GLP1和肽YY,它们在胰岛素分泌中发挥作用。此外,由于微生物BCAA(支链氨基酸)生物合成和BCAA降解的比例改变,BCAA水平升高与人类研究中的早期胰岛素抵抗相关,可能由普雷沃菌(Prevotella copri)和拟杆菌(Bacteroides vulgatus)驱动。P. copri的定植还与常规小鼠模型中的胰岛素抵抗相关。另一种微生物代谢产物,咪唑丙酸,在T2DM患者中升高,可以直接损害葡萄糖耐量和胰岛素信号传导。这些研究突出了微生物群衍生的循环代谢物如何驱动糖尿病的发病机制。
患有2型糖尿病(T2DM)的患者主要通过饮食和药物(如二甲双胍、磺脲类药物和胰高血糖素样肽1(GLP1)激动剂)进行管理,尽管他们最终可能需要胰岛素替代疗法和/或手术。目前的方法是依次尝试治疗选项,尽管不同个体对每种治疗的反应存在很大差异,一些患者可能对饮食或药物没有反应。微生物群为优化提供了一个重要的途径,以确定某种治疗策略是否更适合个体。在以色列进行的一项开创性研究中,对800名非糖尿病个体进行了研究,概述了基于一个优雅的机器学习方法开发个性化饮食建议的潜力,该方法使用包括微生物群和宿主特征以及对不同饮食的血糖反应的一系列测量(见图1;补充图1)。随后的一项研究在美国中西部的327名无糖尿病个体中验证了这种方法,并确认个体的微生物群可以预测对不同餐食的血糖变化。有趣的是,作为一组的碳水化合物仍然与增加的血糖反应相关,但这种方法可以识别碳水化合物中的主要问题个体,允许他们限制特定的碳水化合物,而不是通常遵从性较低的全面低碳水化合物饮食。另一项研究发现,个体的微生物群不仅可以预测血糖变化,还可以预测对不同餐食的甘油三酯变化。
微生物群与糖尿病治疗
治疗糖尿病最常用的药物之一是二甲双胍,它抑制肝脏葡萄糖产生,增加胰岛素敏感性,并增强肌肉和肝脏的葡萄糖摄取。二甲双胍的疗效似乎至少部分依赖于微生物群。在动物和人类的研究中,二甲双胍的使用导致阿克曼氏菌(A. muciniphila)的丰度增加,以及与短链脂肪酸(SCFAs)产生相关的几种细菌种类(例如,Blautia和Butyricicoccus)的增加。阿克曼氏菌可以通过增加肠道杯状细胞的增殖、减少胃肠通透性、降低内毒素血症以及刺激TLR信号传导来改善血糖控制。短链脂肪酸丁酸通过其对骨骼肌、棕色脂肪组织和胰岛β细胞的有益效果,与啮齿动物能量代谢的改善有关。此外,短链脂肪酸丙酸抑制肝脏的糖异生作用,并在啮齿动物模型中减少食欲和体重。二甲双胍最常见的不良反应与胃肠不适有关,如疼痛、膨胀和恶心。包括27名未患糖尿病的健康男性的一项研究发现,在开始二甲双胍治疗之前,粪便中特定菌属(Sutterella、Allisonella、Bacteroides和Paraprevotella)的丰度与胃肠不良反应的发展有关。这一发现表明,除了在二甲双胍疗效中的作用外,肠道微生物群也可能促成其胃肠不耐受。因此,基于微生物群的分层可以使可能有利的反应并能够耐受治疗剂量的患者被筛选出来。支持肠道微生物群在其他糖尿病治疗中作用的数据很少,但在小鼠中使用GLP1激动剂利拉鲁肽后,拟杆菌门(Firmicutes)的丰度减少与血糖控制的改善相关。
鉴于支持二甲双胍使用与阿克曼氏菌(A. muciniphila)丰度增加以及丁酸产生微生物增加之间的联系的数据,一项多中心、双盲、随机安慰剂对照试验研究了在76名2型糖尿病患者中以益生菌形式给予这些微生物的效果。与安慰剂相比,接受组合疗法合生元(A. muciniphila、Clostridium beijerinckii、Clostridium butyricum、Bifidobacterium infantis、Anaerobutyricum hallii和菊粉)的患者血糖控制有向更好的趋势,尽管小样本和短暂的随访时间(12周)使得这种方法是否对2型糖尿病长期有益尚不清楚。扩大这一发现的试验和使用类似针对性微生物群方法进行糖尿病管理的研究,应有助于将这种状况的治疗进一步推进到未来的精准医疗中。
非酒精性脂肪肝病
非酒精性脂肪肝病(NAFLD)是与代谢综合征相关的相当大的发病率,如果不加以控制,可能会发展为肝硬化和终末期肝病。肠道微生物群通过肠-肝轴与肝脏密切相关,微生物产物的解毒是肝脏的重要功能。
微生物群与非酒精性脂肪肝病(NAFLD)的联系
在代谢综合征中观察到的肠道微生物群的改变及其后果,如拟杆菌门(Firmicutes)与杆菌门(Bacteroidetes)比例升高、能量获取能力增加、肠道通透性增加和低度炎症,也在NAFLD中被报道。微生物群在NAFLD中的作用得到了通过将患有NAFLD的人类供体的粪便转移到无菌小鼠后发生脂肪肝的支持。其他涉及NAFLD的微生物群介导的机制包括微生物胆汁酸修饰及其对肝脏法尼酯X受体(FXR)信号的影响、内毒素血症以及尿毒素如甲胺和对甲酚硫酸盐的产生。尽管根据定义,NAFLD与酒精消费无关,但当前的定义并未考虑内源性乙醇的产生。大肠杆菌和来自变形菌门(Proteobacteria)的其他肠杆菌科细菌能够内源性产生乙醇;因此,这些微生物在肠道中的高代谢活性理论上可能导致被认为患有NAFLD的患者的脂肪肝乙醇水平升高。虽然极端情况(被称为自动酿酒综合征)很少见,但微生物产生的长期低水平乙醇可能仍然是一个促成因素。
微生物群与非酒精性脂肪肝病(NAFLD)治疗
二甲双胍,通常用于治疗2型糖尿病,也用于NAFLD的治疗,动物研究支持二甲双胍在此设置中的疗效,人类数据显示肝功能测试有所改善,但组织学反应并未改善。正如前面所概述的,对二甲双胍的成功反应似乎至少部分由肠道微生物群驱动。NAFLD与肠道微生物群之间的机制联系导致研究探索潜在的基于微生物群的治疗。粪便微生物群移植(FMT)在动物研究中显示出前景,初步的人类数据也表明,在FMT之后肝脏脂肪变性和异常肠道通透性有所改善。正在进行的临床试验将更好地阐明这些程序的疗效和安全性。益生菌的施用在改善脂肪肝和肝脏炎症标志物方面也显示出一些前景,然而,这些益生菌配方的组成变化很大,这使得目前对它们的临床推荐变得复杂。在无菌小鼠中,使用来自两位酒精性肝炎患者的粪便对其进行定植,发现针对细胞溶解性粪肠球菌的噬菌体具有治疗益处,目前尚不清楚类似的方法是否对NAFLD也有用。其他专注于减轻微生物群对肝脏毒性来源的新型微生物群治疗看起来很有前景,但仍处于早期阶段。
心血管疾病
心血管疾病(CVD)是美国的主要死亡原因,并在全球范围内持续上升,2005年至2015年间增长了12.5%。据估计,通过改善生活方式和饮食,90%的心血管疾病是可以预防的。心血管疾病已多次与内毒素血症、增加的肠道通透性和低度炎症联系在一起,所有这些都可能由肠道微生物群驱动。
微生物群与心血管疾病(CVD)的联系
肠道微生物群研究领域的一个早期进展是发现代谢物三甲胺-N-氧化物(TMAO)的血浆水平升高是心血管疾病的一个风险因素。TMAO是在肝脏中由黄素单加氧酶作用产生的,使用细菌代谢物三甲胺(TMA)作为底物。TMA来自细菌对胆碱、卵磷脂或左旋肉碱的转化。基于动物研究的TMAO的机制作用表明,它可能是动脉粥样硬化斑块的主要驱动因素,而且高血浆左旋肉碱水平与心血管事件的无事件生存率降低相关,只有当TMAO水平同时增加时,这表明TMAO可能是人类心血管风险的驱动因素。TMAO对CVD的贡献机制包括它对泡沫细胞和内皮细胞、血管炎症、动脉粥样硬化病变、纤维化以及增强的血小板聚集和血栓形成的影响。特定的肠道微生物,包括普罗威登斯菌(Proteus mirabilis)、普罗威登斯菌(Proteus penneri)和大肠埃希菌(Escherichia fergusonii),已知在体外和动物模型中产生TMA。然而,由于菌株间多样性很大,负责TMA产生的基因,即cutC/D或cntA,在人类肠道微生物群中的丰度可能比特定类群的水平具有更大的预测价值。
此外,人类的动脉粥样硬化性心血管疾病也与肠道微生物群的微生物发酵功能能力降低有关,以及与细菌类群肠杆菌科(Enterobacteriaceae)和链球菌属(Streptococcus)的丰度增加有关。其他与降低血压相关的微生物群衍生代谢产物是短链脂肪酸(SCFAs)醋酸和丁酸。SCFA产生菌,如直肠真细菌(Eubacterium rectale)、长链真细菌(Dorea longicatena)、梭状芽胞杆菌(Clostridium clostridioforme)和费卡利杆菌(F. prausnitzii)的丰度减少,与人类心力衰竭的发展有关。饮食是预防和治疗心血管疾病的主要手段之一,由于微生物对饮食成分的代谢在心血管疾病的发病机制中具有机制作用,肠道微生物群可能部分负责饮食干预的有效性。地中海饮食和高纤维饮食似乎对心血管疾病具有保护作用,一项对396名心肌梗死(MI)患者和843名健康个体作为对照的病例对照研究发现,缺乏普雷沃菌(P. copri)与地中海饮食后MI风险降低18%相关,而携带普雷沃菌与地中海饮食后MI风险非显著增加相关。坚持地中海饮食也与增加几种已知能代谢纤维并产生SCFAs的肠道微生物的丰度有关,如费卡利杆菌、真细菌(Eubacterium eligens)和细胞纤维梭菌(Bacteroides cellulosilyticus)。增加纤维摄入的好处可能与纤维降解微生物或其相互作用伙伴增加产生SCFA醋酸有关。醋酸参与调节转录因子Egr1,后者又调节小鼠心脏炎症、纤维化和肥大。此外,丁酸产生菌梭菌属(Clostridium sphenoides)在各种饮食干预前的丰度增加,与肥胖个体胆固醇水平的更大降低有关,也可能与心血管疾病相关。与地中海饮食和高纤维饮食形成对比的是,西方饮食(高摄入脂肪和/或加工肉类、饱和脂肪、盐、糖和精制谷物)与心血管疾病风险增加有关,这可能与肠道微生物如双歧杆菌(Bifidobacterium)和真细菌属(Eubacterium spp.)的丰度减少有关。有趣的是,TMAO前体胆碱、卵磷脂和左旋肉碱在动物蛋白中普遍存在,这是西方饮食的典型组成部分。然而,食用动物蛋白如红肉可能只对携带能够产生TMA或其他代谢产物的微生物的个体子集有害。
微生物群与心血管疾病(CVD)治疗
针对心血管疾病的几种药物治疗的疗效和毒性与肠道微生物群有关。这方面的一个关键例子是Eggerthella lenta中存在心脏糖苷还原酶基因,该基因使洋地黄毒苷(一种通过抑制心肌中的Na+/K+/ATPase来治疗心律失常的重要药物)失活。很可能这种细菌酶活性是由于底物的多样性而不是对洋地黄环境暴露的反应而进化出的过程。这一发现提供了一个例子,说明肠道微生物群的化学多样性如何能够与人类设计药物的代谢产生交流。由于洋地黄具有狭窄的治疗窗口,确定开始治疗前这种细菌代谢途径的存在可以使剂量更精确,并最小化不良反应。有趣的是,负责洋地黄失活的基因在大量精氨酸存在时被抑制,高蛋白饮食在小鼠模型中减少了洋地黄的失活。
他汀类药物通过竞争性抑制HMG-CoA还原酶,是治疗与心血管疾病相关的高脂血症最常用的药物,在美国40至75岁之间的人口中几乎有一半有使用它们的指征。有趣的是,通过测量LDL胆固醇水平的降低,发现个体对他汀类药物的反应存在相当大的变异性。这种变异性可能源于肠道微生物群,因为在个体和动物模型中观察到更强大的治疗反应与更高的肠道微生物多样性相关。此外,在一项研究中,100名总胆固醇水平在160-400 mg/dl之间且LDL反应不同的个体中,高水平的变形菌门与辛伐他汀的效力降低相关。在64名高脂血症患者的人群中,不同他汀类药物罗苏伐他汀的效力也因高水平的蓝藻和拟杆菌门,以及低水平的厚壁菌门和梭菌门而降低。这些研究表明,可以根据个体的肠道微生物群来预测他汀类药物治疗反应。2020年发表的一项研究发现,与肥胖相关的拟杆菌2(Bact2)肠型在接受他汀类药物治疗的患者中不太普遍,这表明他汀类药物具有塑造微生物群的效果。这一发现是否可以用来预测治疗结果并指导未来的治疗选择,还有待观察。
鉴于支持肠道微生物群在心血管疾病(CVD)发病机制中的作用的证据不断积累,目前正在进行的几项临床试验正在研究益生菌在CVD中的作用,这并不令人惊讶。两个例子包括比较抗菌药物利福昔明和益生菌酿酒酵母,以及研究乳酸菌对心力衰竭患者炎症的影响。这些干预性试验为研究基于患者治疗后收集的肠道微生物群的纵向信息的潜在有益的微生物群重组提供了宝贵的机会。基于这些突出的发现,下一代微生物治疗似乎有其位置,它可以推动特定的功能,如醋酸产生或改善屏障功能。另一种方法是开发特定微生物途径的小分子抑制剂,如最近描述的TMA产生酶的抑制剂。这些抑制剂可能实现更精确的治疗干预,并且可以专门针对那些具有高TMAO水平和功能基因水平的患者,这些水平表明具有高TMAO产生能力。
总的来说,认识到肠道微生物群在CVD发病机制和治疗中的作用是一个重要的进展,它为疾病识别、分层和治疗开辟了新的途径(见表1)。
自身免疫或炎症性疾病
类风湿性关节炎
类风湿性关节炎(RA)是一种自身免疫状况,导致关节的慢性炎症。几项研究已经描述了RA患者的肠道微生物群的改变,这些改变随着疾病阶段的不同而变化。
微生物群与类风湿性关节炎(RA)的联系。文献中一个一致的发现是,普雷沃菌属(Prevotella)的成员(Prevotella histicola)与RA疾病的进展以及疾病的改善相关,这突出表明同一属中不同种和/或菌株可能对宿主生理产生不同的影响。因此,重要的是要解决种或菌株水平上的分类学差异,而不是将整个属一概而论地标记为有益或有害。P. copri在RA发病机制中的潜在作用是基于人类和啮齿动物研究的发现。在体外研究和小鼠中,已显示出P. copri增加TH17反应,而TH17反应又与关节炎症性骨侵蚀增加相关。在人类中,已在RA受影响关节的滑液中发现P. copri的16S rDNA。尽管P. copri似乎是RA的一个重要决定因素,但在健康个体中其水平据报道随时间高度可变;因此,需要进行纵向研究,并结合宿主表型的评估,以更好地理解其在RA中的作用。除了肠道微生物群,特定的牙周细菌和牙周病也与人类和关节炎的小鼠模型中RA风险增加相关。牙龈卟啉单胞菌(Porphyromonas gingivalis)和伴放线放线杆菌(Aggregatibacter actinomycetemcomitans)都与抗瓜氨酸化肽的自身抗体增加有关,可能促成RA中的自身免疫反应。
微生物群与类风湿性关节炎(RA)治疗
除了在发病机制中的作用外,肠道微生物群还可能在决定对RA常用药物的反应中发挥作用。这些药物包括改变病情的药物,如甲氨蝶呤和羟氯喹,以及抗炎药物,如磺胺吡啶和非甾体抗炎药(NSAIDs)。宿主因素和遗传学未能为对甲氨蝶呤的反应提供预测模型,但较高的肠道微生物多样性已与甲氨蝶呤治疗相关。在26名未服用药物的新发RA患者中,发现甲氨蝶呤反应者和非反应者之间存在不同的微生物类群及其基因。使用机器学习技术开发的基于微生物群的模型能够以高度的准确性(AUC 0.84)预测21名患者的甲氨蝶呤无反应。这一发现归因于肠道微生物群直接代谢甲氨蝶呤,因为将药物与来自患者的远端肠道微生物群共同孵化后的甲氨蝶呤水平能够预测临床反应。在另一项研究中,甲氨蝶呤治疗对小鼠模型中特定微生物类群和途径的影响导致免疫激活减少,从而减少了疾病活动。这些研究表明,肠道微生物群对甲氨蝶呤的微生物代谢可能在药物的效力中发挥作用,甲氨蝶呤对减少疾病活动的效应本身是由肠道微生物群的调节驱动的。通过竞争性抑制二氢叶酸还原酶来抑制免疫功能的甲氨蝶呤已与肠杆菌目(Enterobacteriales)的丰度减少相关,但目前尚不清楚这一发现对药物反应是否有任何影响。然而,它确实进一步表明甲氨蝶呤影响了肠道微生物群的结构,并且可以进一步探索基于微生物群的甲氨蝶呤反应预测,以指导治疗。肠道微生物群对甲氨蝶呤本身的不同微生物代谢转化为留在细菌细胞内的非活性或不可及形式是一种可能的机制,通过这种机制肠道微生物群改变了甲氨蝶呤的效力。肠道微生物也在甲氨蝶呤的毒性中发挥作用;发现脆弱拟杆菌(Bacteroides fragilis)灌胃对甲氨蝶呤治疗的小鼠192后的肠道粘膜炎有保护作用,这是大约三分之一使用甲氨蝶呤治疗的患者出现的不良反应。通过减少TLR信号传导和CD154表达来抑制免疫激活的羟氯喹的效力已与肠道微生物alpha多样性相关,较高的治疗前多样性有利于更大的效力,但目前尚不清楚是仅仅是较高的微生物多样性还是特定细菌的增加丰度负责这一效应。与甲氨蝶呤一样,目前已有研究发现与TNF抑制剂依那西普相关的肠道微生物群的改变,但在当前研究中仍未定义与效力的关系。
5-氨基水杨酸前药磺胺吡啶通过肠道微生物群的酶促作用乙酰化转化为其活性代谢产物,因此其疗效依赖于肠道微生物群。与非甾体抗炎药(NSAIDs)和对乙酰氨基酚相关的不良事件可能与肠道微生物群有关。细菌β-葡萄糖醛酸酶的活性可能导致NSAIDs的毒性,而这种酶的抑制剂减少了小鼠中NSAID诱导的肠病。原则上,测量β-葡萄糖醛酸酶活性可以帮助识别应该避免使用NSAIDs或适合与特定小分子β-葡萄糖醛酸酶抑制剂联合治疗的个体。某些细菌可以产生对甲酚,它与肝脏中的对乙酰氨基酚竞争酶结合,并可能导致产生肝毒性化合物NAPQI。因此,对甲酚水平可能被用来指导对乙酰氨基酚剂量,以避免肝毒性不良反应。肠道微生物对RA(类风湿性关节炎)发病机制的潜在贡献导致了益生菌作为潜在治疗选择的探索。这些努力主要集中于调节免疫系统以对抗RA中的变化,而不是可能替代缺失的微生物或机制的策略。P. histicola通过抑制抗原特异性的TH17反应和刺激IL-10的转录增加,降低了易感HLA-DQ8小鼠的关节炎发生率和严重程度。在小鼠的胶原诱导性关节炎模型中,给予干酪乳杆菌与CD4+ T细胞产生的促炎分子(IL-1β、IL-2、IL-6、IL-12、IL-17、IFNγ、TNF和COX2)减少相关。在人类中,已研究凝结芽孢杆菌作为45名RA成人的潜在辅助治疗选择,给药导致疼痛和残疾的自我评估改善,并且炎症标志物低于安慰剂。微生物种类对免疫系统的不同效应表明,基于免疫标记的RA亚型可能有助于选择最有可能对特定基于微生物群的治疗有反应的患者。
炎症性肠病
炎症性肠病(IBD)是一种慢性炎症状况,包括克罗恩病和溃疡性结肠炎。IBD亚型在疾病表型、易感性、进展和治疗反应方面的异质性激发了人们尝试超越仅仅基于临床表现的亚型分类。研究的焦点一直放在宿主基因和免疫反应上,但是暴露组和微生物群越来越被认为是IBD中个体间变异的重要决定因素。
微生物群与IBD的联系
肠道微生物群在IBD中的中心作用是基于以下观察:粪便分流后的疾病缓解、胃肠道细菌水平升高区域的更高疾病负担、抗生素治疗后部分患者的改善,以及IBD特有的肠道微生物群组成和功能变化。此外,在将IBD患者的粪便定植小鼠后,观察到与IBD相关的炎症途径(如IL-17)的增强,突出了其在疾病中的作用。
克罗恩病和溃疡性结肠炎均以微生物群落结构的戏剧性变化为特征,最一致的发现是增加了变形菌门(Proteobacteria)的相对丰度,特别是肠杆菌科(Enterobacteriaceae)。特定种的丰度已与克罗恩病(大肠杆菌、弯曲杆菌种和牛型结核分支杆菌)相关,特定丁酸产生菌的耗竭与克罗恩病和溃疡性结肠炎的发病机制相关。在克罗恩病患者的回肠粘膜中也发现了粘附侵袭性大肠杆菌,伴随着TNF分泌的增加,但目前尚不清楚这种细菌是疾病诱导的还是其存在是潜在疾病因素的结果。总体而言,这些群落变化代表了在牺牲专性厌氧菌的情况下,如变形菌门等兼性厌氧菌的增加。这里概述的大多数研究都集中在腔道微生物群上,尽管它在IBD发病机制中很重要,但在识别IBD方面比粘膜相关微生物群具有更低的鉴别力。
IBD中肠杆菌科(Enterobacteriaceae)的总体扩张可能是由于肠道营养景观的变化,例如宿主产生的N-乙酰乙醇胺信号脂质水平升高,这可以被肠杆菌科利用。此外,细菌氮代谢与肠杆菌科的扩张有关,尽管尿素酶的产生和炎症环境中硝酸盐的可用性,这些可以促进肠杆菌科的厌氧呼吸。变形菌门(Proteobacteria)的增加丰度并不一定是IBD中的初始事件,可能是宿主遗传易感性以及饮食和环境暴露的结合结果。然而,变形菌门确实包含高度免疫原性的脂多糖(LPS),其本身就可以触发炎症反应。这种正反馈机制可能有助于持续炎症,并允许变形菌门繁衍生息,同时排除像费卡利杆菌(F. prausnitzii)这样在炎症环境中不适应的细菌。这一假设部分得到了观察的支持,即IBD的治疗与TNF抑制剂与克罗恩病儿童患者肠道微生物群多样性的恢复相关。除了LPS,其他几个细菌组分和代谢产物也与IBD有关,例如多胺和ATP水平更高,次级胆汁酸和丁酸水平更低。
微生物群与IBD治疗
肠道微生物群也在预测对现有IBD治疗的反应中发挥作用。治疗前较高的肠道微生物alpha多样性与使用抗整合素疗法vedolizumab(α4β7拮抗剂)治疗后缓解的可能性更高相关,这表明微生物代谢在决定疗效中可能发挥作用。同样,特定的肠道微生物群特征已与抗TNF治疗药物英夫利昔单抗停用后的疾病复发相关。由于当前的临床实践通常是在实现缓解后很长时间内继续生物制剂治疗,这些特征可能允许选择可以逐渐停药的患者。鉴于对生物药物反应的变异性以及所涉及的成本和发病率,通过微生物群分析预测反应和持续缓解的能力可以简化IBD的管理。这些观察结果促进了基于微生物群治疗IBD的治疗开发,范围从益生菌和粪便微生物群移植(FMT)到特定的细菌化合物或代谢产物。费卡利杆菌(F. prausnitzii)是一个可能的益生菌例子,因为克罗恩病患者术后标本中费卡利杆菌的减少与切除后疾病复发的增加相关。费卡利杆菌已被证明可以通过减少炎症细胞因子的分泌来预防急性结肠炎,这表明它具有抗炎作用,这可能是由于其产生丁酸的能力或通过对免疫系统的独立作用。未来,术后微生物群的分析可能使能够识别那些可能从费卡利杆菌益生菌等治疗策略中受益的个体。同样,IBD中FMT的疗效已与供体微生物群中的特定特征相关。毛螺菌科(Lachnospiraceae)和瘤胃菌属(Ruminococcus)的丰度与反应相关,这表明特定的微生物类群或代谢产物在决定反应中的作用,并为迄今为止在其他FMT试验中看到的响应变异性提供了可能的解释。随着我们对肠道微生物群在IBD中的作用了解的越来越多,基于肠道微生物群的亚型分型可能会更好地对患者进行分层,以预测进展以及对特定治疗的反应,并可能导致开发针对肠道微生物群的个性化治疗策略。
食物过敏
食物过敏很可能是由遗传、饮食和共生微生物群之间复杂的相互作用驱动的。在人类中,已经报道了不同类型食物过敏(如对鸡蛋、花生、大豆、小麦和牛奶过敏)的队列中肠道微生物群的明显变化。然而,研究之间缺乏一致性使得解释这些变化具有挑战性。肠道微生物群的作用得到了以下观察结果的支持:无菌小鼠对食物的过敏性反应敏感,抗生素增加过敏原致敏,并且从健康婴儿移植肠道微生物群可以保护小鼠免受食物过敏。在后者的粪便微生物群移植(FMT)研究中,发现细菌种群Anaerostipes caccae对牛奶食物过敏反应具有保护作用。另一项FMT研究发现,使用细菌Subdoligranulum variabile或拟杆菌属(Bacteroides)菌株的联合体进行的细菌治疗对小鼠的花生过敏具有保护作用。具体来说,Treg细胞途径MyD88–RORγt被发现对保护小鼠免受食物过敏很重要,并被发现在有食物过敏的婴儿中缺乏。Treg细胞亚群是由微生物代谢产物丁酸诱导的,但在这项研究中,丁酸并未被发现是观察到的效应的原因。
除了免疫教育,微生物群可能通过产生或降解影响过敏原的有效剂量。特定菌株的牛奶发酵益生菌,如鼠李糖乳杆菌(Lacticaseibacillus rhamnosus),改善了对牛奶的耐受性,支持了其在过敏原降解中的潜在作用。因此,通过评估微生物群降解食物的能力,我们可能能够预测过敏症的自发缓解或识别可能从基于微生物群的治疗中受益的个体。
特应性反应和哮喘
早期生活中肠道微生物群的破坏,例如与剖宫产和在“清洁”环境中长大(微生物暴露减少)相关的情况,已经与发生特应性和哮喘的风险增加有关。尽管没有一致的人类微生物群标志与特应性和哮喘相关,但微生物代谢产物可能直接参与疾病的发生。来自肠道的短链脂肪酸(SCFA)丙酸的循环水平被发现可以通过依赖自由脂肪酸受体3(FFAR3)和树突状细胞功能的方式减少小鼠肺部的炎症。微生物代谢产物可能特别涉及TH2的重编程和过度激活,尽管并非所有哮喘患者都有TH2水平升高。通过一系列代谢组学和微生物遗传学研究,鉴定出参与哮喘发展的微生物代谢产物——亚油酸衍生物12,13-二羟基二十碳烯酸(12,13-diHOME)。发现具有增加哮喘风险的出生队列的粪便中12,13-diHOME水平增加,这与肺部抗炎细胞因子和Treg细胞水平降低有关,表明免疫耐受受到阻碍。负责产生12,13-diHOME的细菌环氧化物水合酶可能作为治疗策略,在酶水平升高的患者亚群中被抑制。
已经尝试通过向母乳或配方奶中添加特定菌株或群落,以减少以后生活中发展特应性或哮喘的风险。其中一个例子是使用鼠李糖乳杆菌(L. rhamnosus GG),这增加了被认为促进高风险婴儿发展为哮喘的免疫耐受的微生物种,尽管后来生活中哮喘发展和疾病严重程度的进展没有得到明确评估。此外,从农场牛棚中分离出的嗜麦芽窄食单胞菌(Acinetobacter lwoffii)和乳酸乳球菌(Lactococcus lactis)菌株在小鼠中具有强大的过敏保护特性,可以在人类队列中探索。为了更好地理解肠道微生物群在过敏性和特应性疾病发展中的作用的时间机制,需要追踪临床结果的纵向出生队列。总体而言,这些研究可以为基于肠道微生物群的过敏性疾病个性化治疗策略的未来努力提供信息。
未来展望
在考虑微生物群在健康和疾病中的作用时,人们非常关注因果关系,但很明显,即使肠道微生物群不是诱发因素,它也可能促成疾病。实际上,肠道微生物群很少是疾病的唯一驱动因素,需要在涉及宿主遗传学、宿主生理反应和环境的系统生物学背景下考虑。我们需要了解肠道微生物群在使个体倾向于疾病状态的复杂调控框架中的位置。
尽管肠道微生物群已与许多疾病相关联,但与其他变量(如宿主(表)遗传学、蛋白质组或转录组)相比,很难量化这种贡献的相对量。在界定不同宿主和微生物群因素的贡献方面面临的一个主要挑战是,很难将宿主和环境因素对微生物群的影响与它们对宿主生物学的影响(独立于微生物群)分开。Vujkovic-Cvijin等人调查了一系列可能由于个体间的巨大变异性而使微生物群相关研究混淆的生活方式、生理和饮食因素。像这样的研究不仅有助于进一步了解微生物群与健康和疾病的关系,还有助于提高未来工作的质量,以更好地理解微生物群在精准医学和个体化治疗中的力量。在定义的模型系统中采用多组学方法,如无菌动物,也有助于从不同数据层解开这些贡献。
未来在大量表现型良好的患者队列中进行的研究将需要是多维的,结合宿主和微生物的多组学数据以及暴露组,以更好地理解肠道微生物群和其他数据层面对疾病状态或治疗效果的相对贡献。这些研究还应该通过使用纵向抽样来包含时间和个体间变异。纵向数据分析已被证明与横断面研究相比可以减少变异,并可以用来建立因果关系。例如,通过纵向监测未接受治疗的队列,可以识别出潜在的治疗反应变化背后的相关微生物群和宿主因素,从而识别出治疗反应的基础变异。这种方法结合智能的体外和离体方法可以识别出负责治疗反应、不良反应和药物代谢的新因素。这样的研究还将使我们能够确定肠道微生物群是否可能作为饮食、遗传、年龄和生活方式等其他宿主相关因素的读数,并可能简化疾病分类和治疗分层的机器学习算法(见图3)。
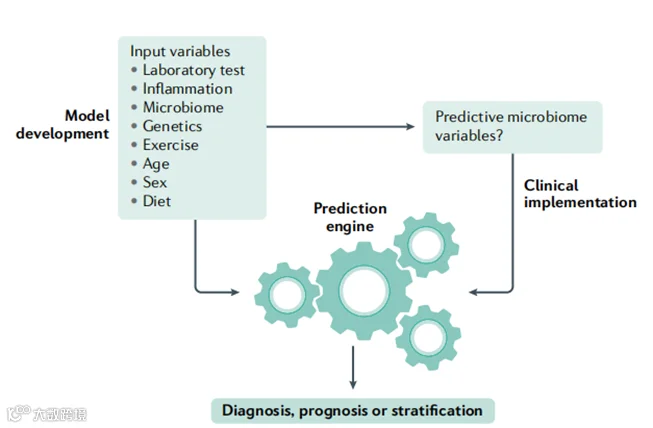
图3 | 肠道微生物群作为预测模型中宿主变量的读数。
使用人工智能和机器学习构建的计算预测模型对优化临床管理具有巨大的前景。这些模型的预测价值取决于训练数据,理想情况下包括一系列宿主测量(左图)。然而,确定每个患者的所有这些参数可能在成本和时间上是禁止性的。由于肠道微生物群对大多数生活方式因素都有反应,它可以作为一个有价值的抽象读数,便于访问和测量。在全面的大规模训练队列(左)中可以识别出最相关的微生物群特征,允许单独使用微生物群作为临床结果的预测器。
正如已经提到的,将整个属描述为有益或有害可能是误导性的,当分析个体微生物群时,应使用更高分辨率的技术。这种二分法的一个好例子是普雷沃菌属(Prevotella),它在类风湿性关节炎(RA)中既被称为“毒药”也被称为“灵丹妙药”。鉴于微生物代谢产物水平与疾病之间的关联通常比分类学丰度更强,微生物功能可能是更相关的靶标。此外,微生物群的功能特性在个体间更为保守。因此,与流行的“缺失微生物”概念相比,“缺失微生物功能”可能是一个对精准治疗更有用的概念。同时,重要的是要注意,一个疾病状态可能有多个微生物功能作为基础,如对艰难梭状芽胞杆菌感染(CDI)的易感性所示。因此,未来的治疗需要基于更好的机制理解,并针对一个或多个缺失的功能,意识到不同个体可能存在不同的功能异常。
随着该领域持续强劲的势头和越来越多的研究,我们可能会在能够稳步前进之前就急于求成。我们需要小心,并意识到我们仍处于研究的早期阶段,在将微生物群的知识转化为可用于临床以使患者受益的可行措施之前,仍有重大挑战需要解决。这些挑战包括在大型临床队列中缺乏对基于微生物群的标记的验证、处理和分析微生物群数据的标准化缺乏,以及对不同疾病状态下与微生物群相关的机制缺乏个性化理解。尽管存在这些不足和挑战,该领域的快速发展和过去十年所取得的进展带来了乐观,即微生物群将很快成为临床实践的一部分。我们可以开始设想如何将微生物群纳入个性化治疗策略,并超越对所有患者采用一系列治疗策略的方法,转而为每个患者选择最佳策略。
结 论
在这篇综述中,我们强调了对肠道微生物群在不同疾病状态的发展、进展和治疗中作用的理解方面取得的主要进展。总体而言,这些证据清楚地突出了肠道微生物群在精准医学和个体化治疗中的前景。虽然已经取得了重大进展,但科学研究的严谨性和支持肠道微生物群作用的发现的强度因疾病状态而异,如本综述所总结(见图1;补充图1)。尽管大多数人类研究都集中在组成分析上,以识别反应的微生物生物标志物,但要发展精准治疗和分层治疗,将需要更深入的机制理解。总体而言,该领域已经意识到这一需求,并已转向研究微生物群的功能方面。尽管非靶向的基于生态系统的方法在CDI(艰难梭状芽胞杆菌感染)中显示出益处,但这种非靶向措施的效力可能不会扩展到多因素的慢性疾病,在这些疾病中,有多种机制在起作用。
往期推荐

