

微生物群影响代谢健康,但特异性微生物分子及作用机制仍有待阐明。作者从可促进小鼠瘦体质的产芽孢菌群中分离出一株Turicibacter菌株。人类宏基因组分析显示,肥胖个体的Turicibacter丰度降低。类似地,高脂饮食会降低Turicibacter定植,阻断其抑制体重增长的作用,但该效应可通过持续补充Turicibacter来逆转。高脂饮食导致神经酰胺蓄积,进而促进体重增加。转录组学和脂质组学分析显示,该产芽孢菌群及Turicibacter可抑制宿主神经酰胺。Turicibacter能够产生独特的脂质分子,而高脂饮食会降低其水平。这些脂质可被转移至宿主上皮细胞,减少神经酰胺生成并降低脂肪吸收。使用纯化的Turicibacter脂质处理动物可预防肥胖,这表明细菌脂质能够促进宿主代谢健康。这些数据鉴定出细菌与宿主之间的一个脂质代谢环路,该环路可被饮食破坏,并可作为治疗靶点。
1、 产芽孢菌群中的单个成员Turicibacter可改善代谢健康
Turicibacter KKT8被鉴定为一种能够减少脂肪积累和血清甘油三酯的产芽孢菌群成员。
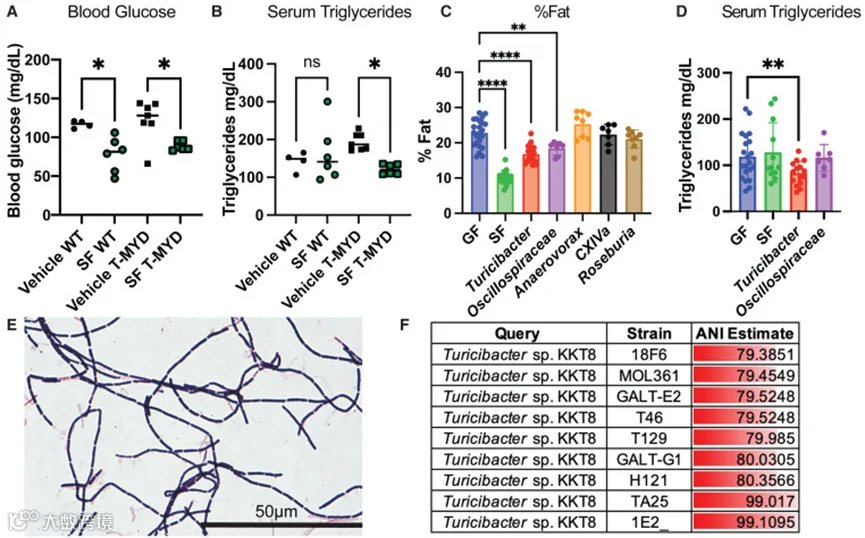
图1. 产芽孢菌群中诱导瘦体质微生物的鉴定
2、 产芽孢菌群及Turicibacter可下调小肠和血清中的鞘脂代谢
作者之前已表明产芽孢菌群可下调脂肪酸摄取,但其机制尚不明确。为阐明产芽孢菌群如何影响肠上皮细胞(IEC)生物学,采用激光捕获显微切割(LCM)技术从无菌(GF)和产芽孢菌群定植小鼠的小肠十二指肠(SID)中分离绒毛和隐窝细胞。产芽孢菌群定植后,基因表达发生广泛改变。靶向代谢通路分析显示,与无菌和无特定病原体(SPF)小鼠相比,产芽孢菌群小鼠中许多参与鞘脂代谢(包括神经酰胺代谢)的基因被下调(图2A、2B)。
血清神经酰胺是心血管疾病的预测因子,而心血管疾病是肥胖人群的重要合并症和死亡原因。值得注意的是,Turicibacter KKT8或SF菌群定植可降低血清神经酰胺(图2N和2O)。Turicibacter KKT8单菌定植降低了多种血清脂质,包括二氢神经酰胺、神经酰胺、鞘磷脂、植物神经酰胺、己糖基神经酰胺和磷脂酰胆碱(图2P)。血清脂质的大量下调表明Turicibacter KKT8能够影响肠道和全身性脂质代谢。这些数据支持了此前对Turicibacter sp.的观察结果,并扩展了Turicibacter sp.对宿主血清脂质组的影响。总而言之,这些数据确定,来自复杂产芽孢菌群的一种细菌——Turicibacter KKT8,能够导致瘦弱体质、降低血清甘油三酯并抑制鞘脂代谢,揭示了其在代谢健康中的关键作用。
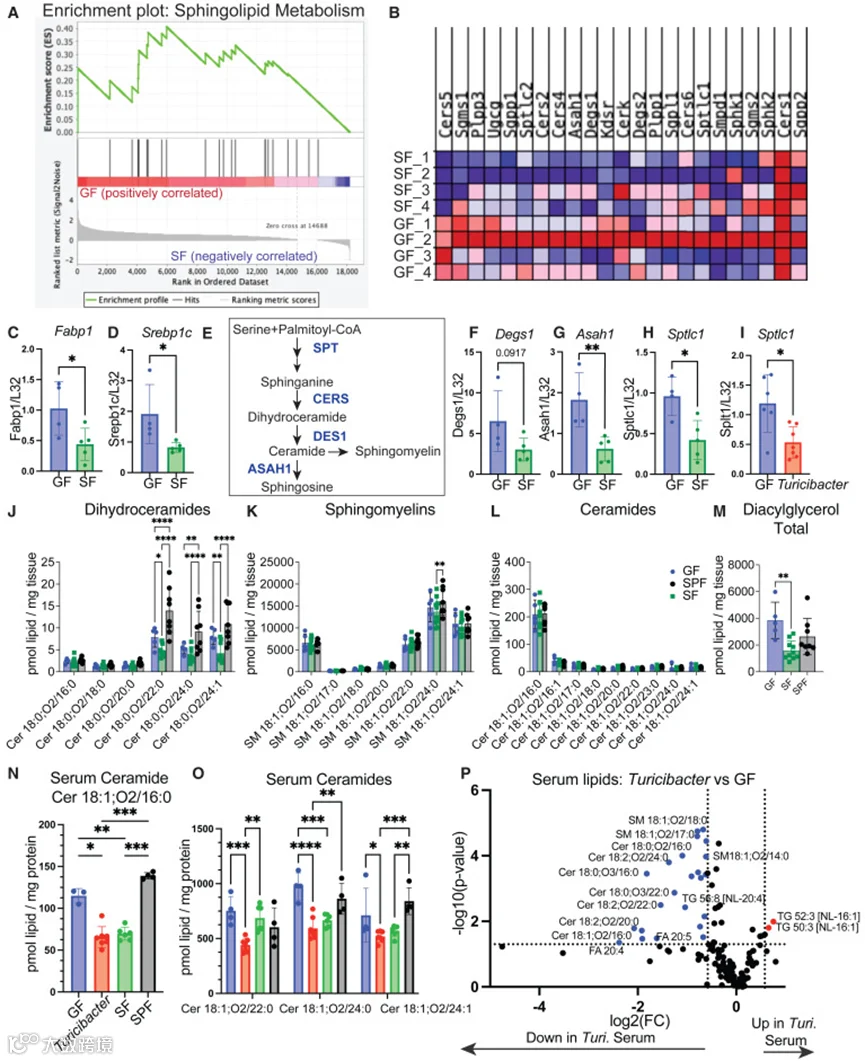
图2 产芽孢菌群和Turicibacter降低肠道和血清中的鞘脂
3、 高脂饮食损害Turicibacter的定植和生长
作者的研究发现Turicibacter KKT8能够促进小鼠的瘦体质和代谢健康,这促使作者研究肥胖人群中Turicibacter的水平。使用GMrepo curated human metagenomic database,作者发现肥胖个体的Turicibacter水平显著降低(图3A),这与其它几项研究一致。综上所述,这些数据表明Turicibacter定植丰度的降低与肥胖个体相关。
为了验证这一点,将Turicibacter KKT8单菌定植动物置于NC或HFD饮食条件下,并沿肠道分析了Turicibacter KKT8的丰度。引人注目的是,尽管它是肠道内唯一的微生物,HFD几乎将Turicibacter KKT8从小肠(SI)中清除(图3C–3E),并显著降低了其在下消化道(GI)中的丰度(图3F和3G)。这些数据证明,高脂饮食可以直接降低Turicibacter的定植。
棕榈酸(PA)是鞘脂代谢的基本构建单元,也是高脂饮食的主要饱和脂肪成分。为了确定PA是否影响Turicibacter KKT8的生长,作者在递增浓度的PA中培养Turicibacter KKT8(图3H)。与对照相比,含PA培养基中的生长显著降低(图3H),当PA添加到活跃培养物中时,生长停止(图3I)。PA对Turicibacter KKT8不一定具有毒性,因为将细菌从含PA的培养基中移出后,其能够恢复生长(图3J)。
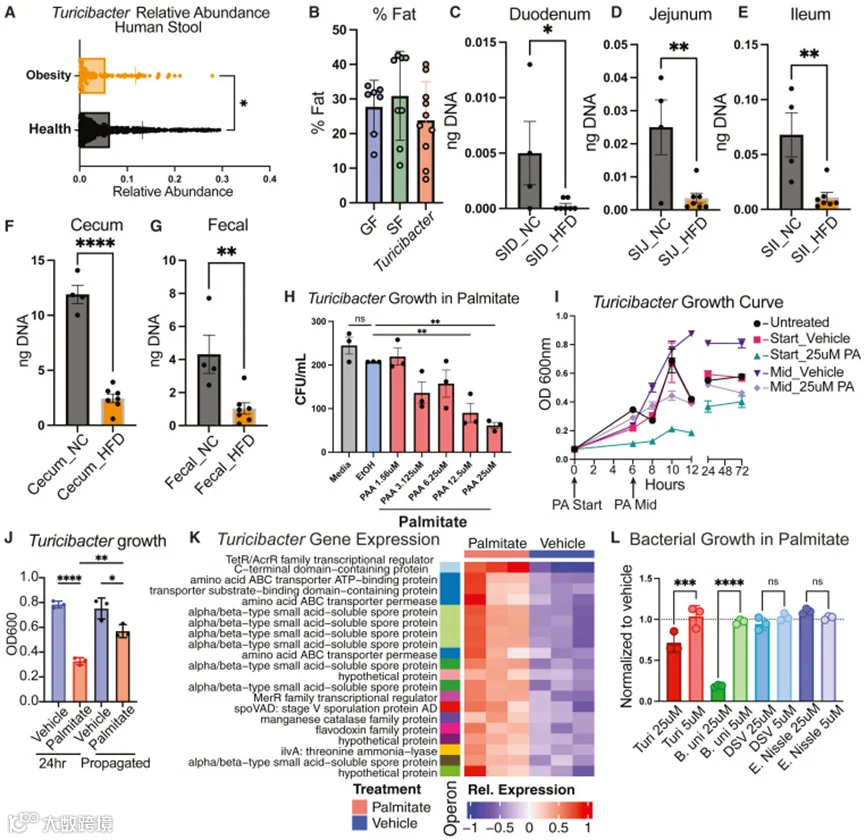
图3 脂质损害Turicibacter生长
4、 持续补充Turicibacter KKT8可在高脂饮食期间预防肥胖并降低宿主神经酰胺水平
T-MyD88−/−小鼠接受高脂饮食,并每周5天口服Turicibacter KKT8。接受补充的动物体重增加减少,体脂、腹股沟白色脂肪组织(iWAT)、空腹血糖和血清神经酰胺均降低(图4A–4F)。因此,在会发生菌群依赖性肥胖的T-MyD88−/−遗传模型中,口服补充Turicibacter KKT8能够减缓高脂饮食相关的体重增加和代谢变化。同样,接受高脂饮食的WT SPF小鼠也能从Turicibacter KKT8中获得代谢益处,表现为体重增加减少、iWAT质量降低以及多种血清脂质(包括甘油三酯和神经酰胺)水平下降(图4G–4L)。高脂饮食下的WT动物补充Turicibacter KKT8还降低了十二指肠中的多种脂质,包括神经酰胺、鞘氨醇和鞘磷脂,以及多种脂肪酸和磷脂酰胆碱。
这些结果突显Turicibacter KKT8补充可抑制高脂饮食的不良代谢效应,从而为肥胖和代谢疾病患者提供了一种潜在的治疗干预策略。
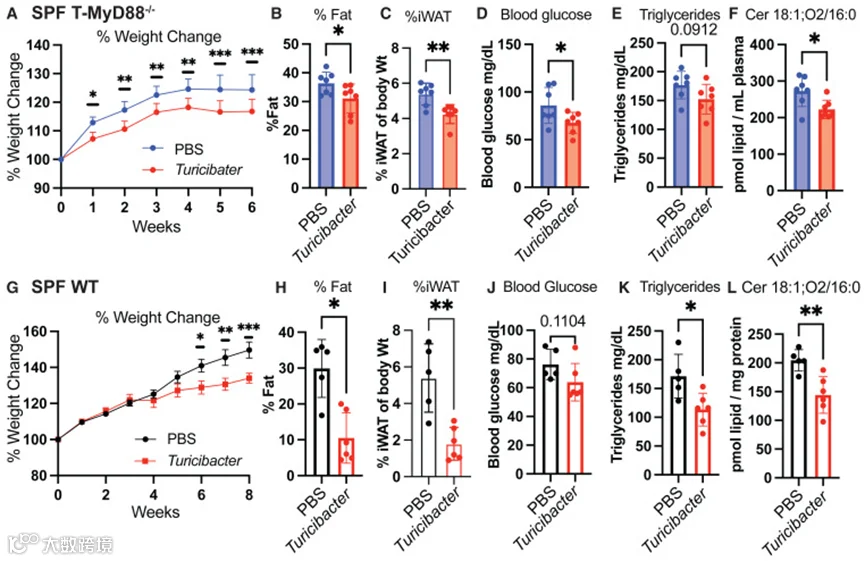
图4 持续补充Turicibacter在高脂饮食下的代谢保护作用
5、 膳食脂质可调节Turicibacter的脂质代谢
高脂饮食会负面影响Turicibacter的生长及其代谢保护能力,而其定植或补充会影响宿主脂质和鞘脂代谢,这表明脂质代谢在Turicibacter KKT8生物学中起关键作用。
为探究Turicibacter KKT8是否具有类鞘脂合成蛋白,作者检测了细菌生长是否对选择性抑制SPT(myriocin)、神经酰胺合酶(fumonisin B1)和DES1(fenretinide)的药物产生响应(图5G)。尽管PA(棕榈酸)显著抑制Turicibacter KKT8的生长,但添加这三种抑制剂中的任何一种均可逆转PA诱导的生长缺陷(图5H–5J)。myriocin和fumonisin B1均为真菌产生的天然产物,对其靶点具有高度特异性,且结构与鞘脂相似。有趣的是,喂食fumonisin B1的小鼠其肠道菌群中Turicibacter丰度增加。由于这些对鞘脂合成酶具有高特异性的鞘脂抑制剂能够影响Turicibacter KKT8的生长,提示Turicibacter KKT8所产生的酶和脂质与鞘脂具有足够的结构同源性,从而允许这些鞘脂抑制剂结合。此外,Turicibacter KKT8还能摄取鞘氨醇(图5K)。利用鞘氨醇的能力主要局限于拟杆菌属细菌,该类菌已知可合成鞘脂。此前已鉴定Turicibacter具有摄取鞘氨醇的能力。综合这些数据表明,Turicibacter可产生多种已知脂质(包括脂肪酸)以及许多尚未被注释的未知脂质,这些脂质中某些可能在结构上与鞘脂相似。未来的研究将对这些脂质进行深入分析。
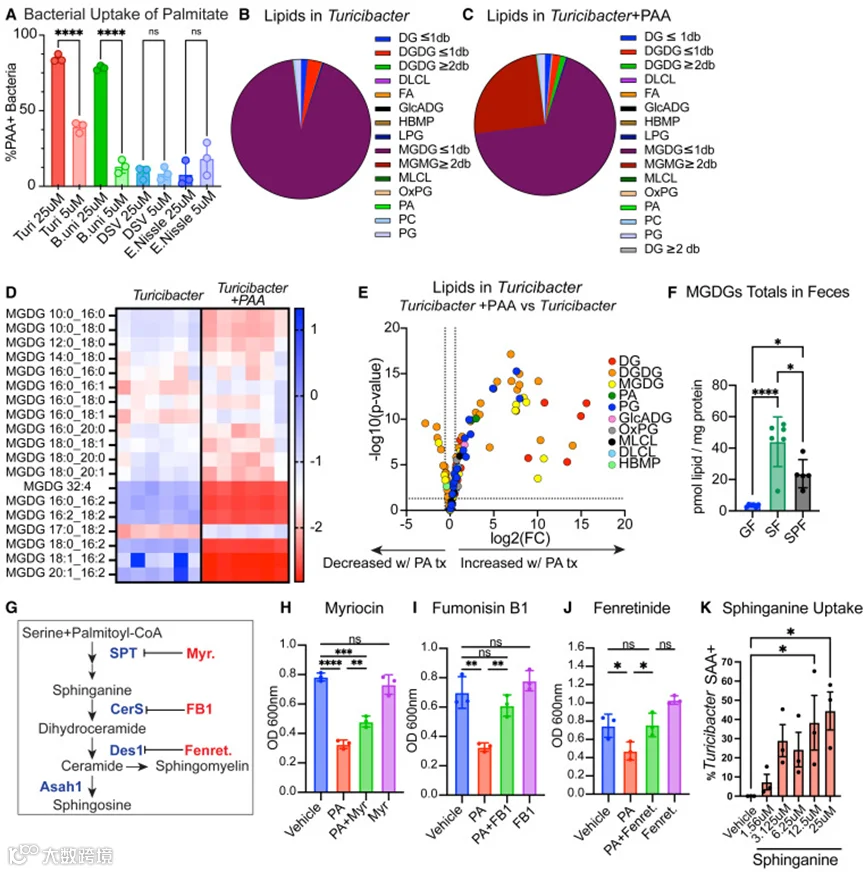
图5 膳食脂质调节Turicibacter脂质代谢
6、 Turicibacter脂质进入肠上皮细胞,调节宿主神经酰胺代谢
拟杆菌脂质已被证明可转移至宿主细胞并影响代谢。鉴于Turicibacter KKT8可产生多种脂质,作者检测了其脂质是否同样可以转移至宿主细胞。将用PAA培养的Turicibacter KKT8施用于小鼠小肠细胞系(MODE-K细胞)。利用点击化学对携带Turicibacter KKT8来源脂质的MODE-K细胞进行荧光标记,并通过流式细胞术进行定量(图6A和6B)。超过40%的MODE-K细胞具有细菌标记的脂质(图6C),提示宿主上皮细胞摄取了Turicibacter KKT8产生的脂质。
与体内研究结果一致,Turicibacter KKT8暴露降低了MODE-K细胞中Srebp1c和Sptlc1的表达(图6D和6E)。对Turicibacter KKT8处理的MODE-K细胞进行脂质组学和脂质本体(LION)分析发现,脂质储存、甘油三酯、鞘脂和神经酰胺均显著下调(图6F–6H),这与作者关于Turicibacter KKT8单定植或补充可降低血清甘油三酯和神经酰胺的发现一致(图1D、2N、2O、4F、4K和4L)。这些数据表明Turicibacter KKT8可将脂质递送至宿主上皮,并且这与降低促进宿主肥胖的神经酰胺相关。此外,膳食脂肪的存在可调控Turicibacter KKT8的脂质代谢,从而阻止特定宿主神经酰胺的下调。
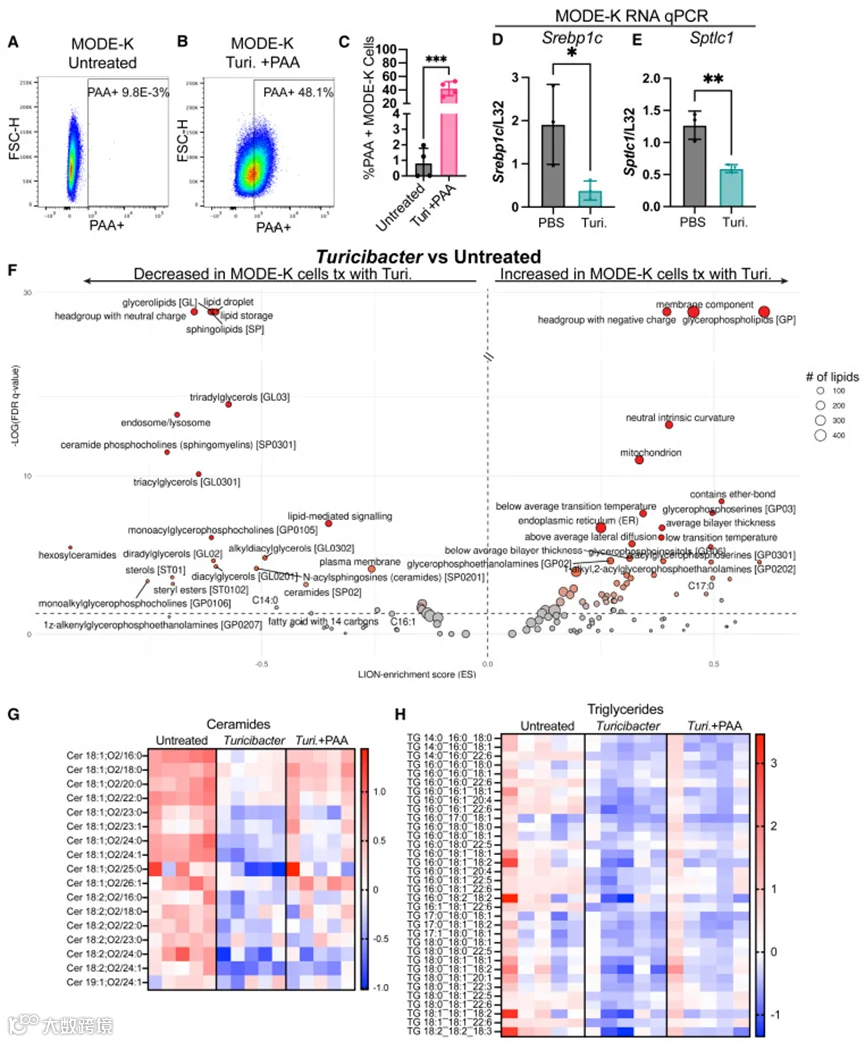
图6 Turicibacter脂质进入上皮细胞并在体外下调神经酰胺和脂质代谢
7、 Turicibacter脂质可减少肠上皮细胞对脂肪酸的摄取并预防肥胖
作者的数据表明,Turicibacter KKT8 能产生独特的脂质,这些脂质可转移至上皮细胞并下调与肥胖相关的宿主神经酰胺水平。此前作者已证实,产芽孢菌群可降低肠道脂肪酸摄取,而其他研究则表明神经酰胺通过 CD36 促进摄取。综合这些研究提示,Turicibacter KKT8 的脂质可能介导了其促进瘦体质的效应。为验证这一点,作者测定了经 Turicibacter KKT8 细菌、Turicibacter KKT8 脂质或其相应溶媒对照处理后的 MODE-K 细胞的脂肪酸摄取情况。正如假设,用 Turicibacter KKT8 或 Turicibacter KKT8 脂质处理 MODE-K 细胞可显著降低脂肪酸摄取(图 7A)。Turicibacter KKT8 的条件培养基(CM)同样减少了 MODE-K 细胞的脂肪酸摄取(图 S7),提示 Turicibacter KKT8 中的生物活性分子为分泌型分子。此外,与完整 Turicibacter KKT8 处理相似,Turicibacter KKT8 来源的脂质处理也可降低 MODE-K 细胞的总神经酰胺水平,效果类似于用 SPT 抑制剂 myriocin 处理的细胞(图 7B)。
为评估Turicibacter KKT8脂质的体内效应,将T-MyD88−/−小鼠饲喂高脂饮食并用Turicibacter KKT8脂质进行灌胃。4周后,经Turicibacter KKT8脂质处理的动物体重增长显著减少,空腹血糖降低,且腹股沟脂肪垫呈缩小趋势(图7C–7E)。此外,Turicibacter KKT8脂质处理显著降低了小肠十二指肠Sptlc1和CD36的表达(图7F和7G),与神经酰胺通路下调一致。这些数据共同表明,Turicibacter通过产生可下调宿主神经酰胺生成的生物活性脂质来促进代谢健康并预防肥胖。
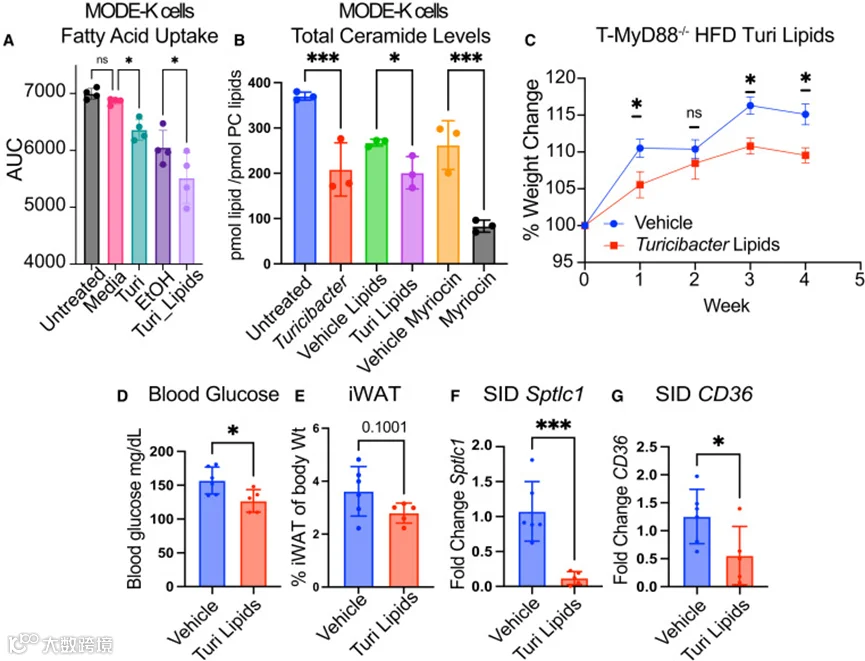
图7 Turicibacter脂质可减少肠上皮细胞对脂肪酸的摄取并预防肥胖
膳食显著影响微生物群组成,但对特定成分如何影响单个生物体及其功能却知之甚少。尽管多种代谢物来源于微生物,但只有少数如短链脂肪酸和次级胆汁酸得到了充分表征。在此作者鉴定出Turicibacter KKT8,这是一种在高脂饮食和肥胖个体中减少的微生物群成员,其产生的独特脂质能够预防哺乳动物宿主的肥胖,揭示了一类具有代谢益处的新型分子。本研究中作者发现,将产芽孢菌群或Turicibacter KKT8添加到缺乏这些微生物的T-MyD88−/−小鼠中,比已拥有这些微生物的野生型小鼠产生更大益处。Turicibacter丰度与膳食脂肪和肥胖呈负相关,提示其作为生物标志物的潜力。然而,Turicibacter水平升高与帕金森病和抑郁症等疾病相关,表明存在菌株水平效应。理解Turicibacter在种水平和菌株水平上的变异如何影响其对健康与疾病的调控作用,对于开发其治疗潜力至关重要。通过精准医学方法,确定哪些Turicibacter缺乏且患有代谢疾病的个体,可能揭示出最能从基于Turicibacter的疗法中获益的患者群体。总体而言,本研究的数据鉴定出一个促进宿主代谢健康且具有治疗前景的新型细菌-宿主脂质网络。
参考文献:
Kendra Klag, Darci Ott, Trevor S Tippetts et al. Dietary fat disrupts a commensal-host lipid network that promotes metabolic health. Cell Metab. 2025 Nov 5: S1550-4131(25) 00441-3. doi: 10.1016/j.cmet.2025.10.007.
