
奇辉观点
暴露于环境的污染物是肿瘤发展的重要易感因素。与药物分子类似,污染物通常在体内被代谢,这可以改变它们的致癌潜力,并通过改变毒代动力学影响组织分布。尽管最近的研究表明,与人类相关的微生物可以化学转化广泛的外源性物质,影响其产生的代谢物并导致组织暴露,但微生物生物转化对化学诱导的肿瘤发展的影响仍然不清楚。本文作者展示了肠道微生物群的耗竭影响亚硝胺的毒代动力学,显著减少了小鼠亚硝胺诱导的尿膀胱癌的发展和严重程度。作者通过使用个体化的细菌培养集合和无菌小鼠模型,分别在体外和体内将这种致癌物生物转化与特定的肠道细菌分离株联系起来,同时测试了来自不同人类供体的肠道群落,以证明微生物致癌物代谢在个体之间存在差异,并且证明这种代谢活性适用于结构相关的亚硝胺致癌物。总的来说,这些结果表明,肠道微生物群对致癌物的代谢可能是化学诱导的致癌作用的一个促成因素,这不仅为理解亚硝胺致癌物的代谢机制提供了新的视角,也为未来的研究奠定了基础,在鉴定微生物组编码的易感风险因素上,并可能实现针对个体微生物群的癌症预防策略。
论文ID
本文译自:Roje B, Zhang B, Mastrorilli E, et al. Gut microbiota carcinogen metabolism causes distal tissue tumours. Nature . 2024 Jul 31. doi: 10.1038/s 41586-024- 07754-w.
发表杂志:Nature
影响因子:49.7
通讯作者:Michael Zimmermann
作者单位:University of Heidelberg
引 言
越来越多的证据表明,与人类相关的微生物群落,统称为人类微生物组,在癌症的发展和进程中扮演着重要角色。尽管微生物对癌症发展贡献的分子机制在很大程度上仍然未知,但已经证明,从肠道迁移到组织中的肿瘤诱导细菌和微生物产生的基因毒素(例如colibactin)可以诱导癌症发展。另一个重要的致癌机制是暴露于环境污染物,这些污染物进入人体后通常会进一步被代谢。已广泛描述了污染物代谢的人类酶,如细胞色素P450酶,它们有能力转化多种致癌物,这可能导致它们的激活或失活。近期的研究表明,人类肠道细菌也有潜力代谢化学多样性化合物,如医疗药物,产生的结果可以分布到远端组织。这使我们去研究肠道微生物组是否可以通过代谢环境致癌物来促进肿瘤的发生和发展。
我们以膀胱癌为例进行了研究,它是全球第十大常见癌症,在侵袭性形式中具有高复发率和进展率,其主要风险因素是环境和职业暴露于致癌物。诸如N-丁基-N-(4-羟基丁基)-亚硝胺(BBN)之类的亚硝胺化合物和烟草烟雾中的相关有机污染物已被充分证明可诱发膀胱癌。事实上,慢性暴露于BBN通常被用来在啮齿动物模型中诱发膀胱癌。口服给药后,BBN通过氧化和葡萄糖醛酸化途径经历大量代谢。值得注意的是,N-丁基-N-(3-羧丙基)-亚硝胺(BCPN),BBN的氧化产物,通过在尿路上皮形成DNA加合物诱导肿瘤形成。我们利用BBN诱导的膀胱癌小鼠模型来研究肠道微生物群对化学诱导的肿瘤发生的贡献以及其在毒代动力学中的作用(图1a)。
结 果
抗生素减少膀胱肿瘤
为了测试肠道微生物群对BBN诱导的膀胱癌发展可能的贡献,我们比较了同时通过饮水(体积比0.05%)给予BBN的经抗生素(ABX)处理和未处理的C57BL/6小鼠的膀胱肿瘤病理。我们通过菌落形成单位计数确认了ABX处理后肠道细菌负荷的减少,在处理过的动物中比未处理的动物低99.99%。此外,16S rRNA扩增子测序表明,ABX处理(而非BBN给药)显著降低了肠道微生物组的α-多样性,并显著改变了整体群落组成。在连续12周暴露于饮水中的BBN后,移除BBN 8周,并对小鼠的膀胱进行了盲法病理分析(图1b)。值得注意的是,大多数肠道细菌负荷减少的小鼠(ABX-BBN组)完全没有膀胱肿瘤病理,而没有ABX处理的对照小鼠中有77%(BBN组)在膀胱组织中发展了肿瘤变化。此外,侵袭性肿瘤,即最具有侵略性的肿瘤表现,在BBN组的小鼠中占53%,而在ABX-BBN组中仅占12%。这些结果表明,肠道微生物群可能会促进BBN暴露小鼠的膀胱癌发展。
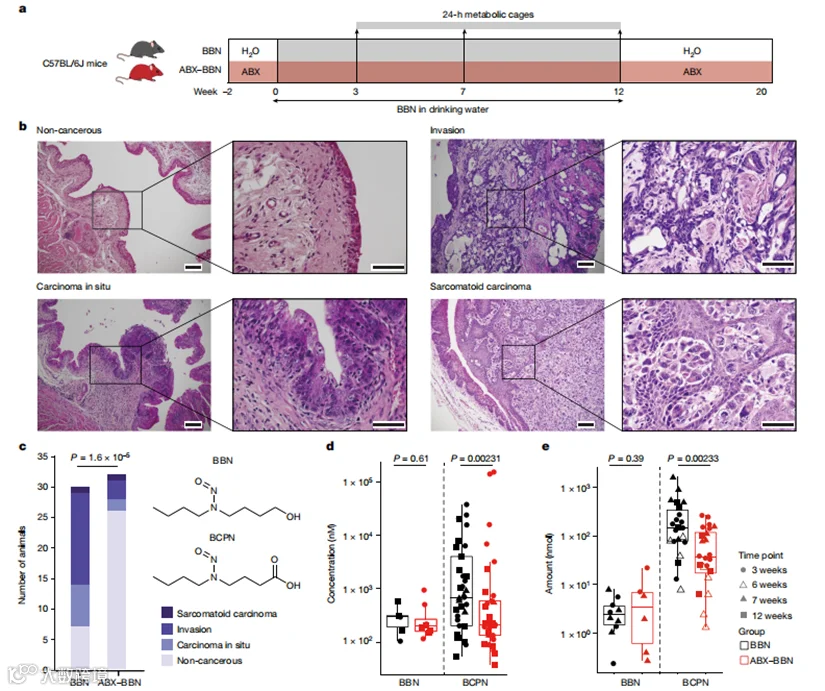
图1 :ABX处理减少了小鼠的化学致癌作用
BCPN是在肠道产生的
为了阐明肠道微生物群与BCPN产生之间的可能联系,我们在BBN暴露3周和7周后,对BBN处理和ABX-BBN处理动物的九个额外体腔(血浆、肝脏、肾脏、十二指肠、空肠、回肠、盲肠、结肠和直肠)中的BCPN进行了定量(图1a)。虽然我们在两组小鼠的血浆、肝脏和肾脏中未检测到BCPN浓度的任何差异(图2a),但在BBN处理的动物(而非ABX-BBN处理的动物)的盲肠和结肠腔内容物中BCPN积累(图2b)。我们还观察到BCPN水平从盲肠到直肠逐渐降低,这表明BCPN从小肠重新吸收。为了控制ABX对宿主BBN代谢的影响,我们用饮水中BBN处理无菌小鼠10天,有无ABX处理,并比较了膀胱、肝脏、肾脏、血浆和盲肠中的BCPN水平。所有这些组织中的BCPN水平无论是否经ABX处理都是可比的(扩展数据图2d和补充表15),表明BBN的代谢和毒代动力学不受ABX的影响。为了直接测试ABX对宿主BBN代谢的潜在影响,我们在HEP-G2细胞中测量了肝脏BBN到BCPN的转化,发现它不受ABX添加的影响(扩展数据图2e和补充表16)。这些对照实验巩固了我们的假设,即BBN和ABX-BBN处理动物之间BCPN水平的差异不是由ABX引起的,而是BCPN以微生物组依赖的方式在下肠道产生。
接下来,我们研究了在盲肠观察到的BCPN产生是否由肠道BBN氧化所致。实际上,在经ABX处理的动物中,下肠道的BBN浓度比ABX未处理动物低了一个数量级以上(图2c),这表明游离BBN只能解释肠道BCPN产生的一小部分,其他BBN衍生物也对肠道BCPN产生有所贡献。的确,非靶向代谢组学分析在ABX处理动物的大肠中检测到了葡萄糖醛酸化BBN(gBBN),而在未经ABX处理的动物中未检测到(图2d)。这些数据表明,普遍存在于肠道细菌中的β-葡萄糖醛酸酶将gBBN脱结合后,释放的BBN氧化成BCPN。为了验证这一假设,我们对BBN处理和ABX-BBN处理的小鼠在体外脱糖基化后,对其盲肠gBBN进行了定量,如前所述。的确,与单独用BBN处理的小鼠相比,ABX和BBN联合处理的小鼠盲肠内容物在实验后释放出更多的BBN(图2e和补充表8)。对总BBN(游离BBN和gBBN)和BCPN的比较显示,BBN处理动物体内的BCPN量与ABX-BBN处理动物体内的BBN量相当(图2f和补充表8和11),表明肠道微生物群通过脱糖基化后,从游离BBN和gBBN产生BCPN。
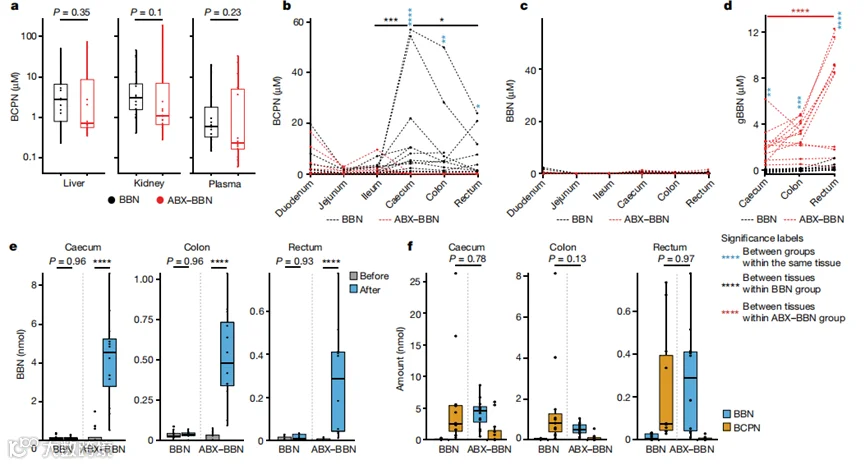
图2 :BCPN是从BBN及其衍生物在大肠中生成的。
肠道细菌将BBN氧化为BCPN
在发现观察到的微生物组依赖性BCPN差异是由gBBN的肠道脱结合和随后BBN的代谢引起的之后,我们试图测试肠道微生物是否可以直接将BBN氧化为BCPN。为此,我们在厌氧、微氧(10%氧气)和好氧(21%氧气)条件下培养了小鼠的盲肠、结肠和直肠样本,并将这些体外微生物群培养物与BBN共孵化。在BBN暴露4、8、12和24小时后对BCPN的定量表明,这些微生物群仅在好氧和微氧气氛下具有将BBN氧化为BCPN的能力,尽管所有条件下的最终细菌密度相当(图3a)。尽管BBN转化为BCPN对氧气的依赖性在反应的氧化性质上并不出乎意料,但我们也在小鼠的大肠中观察到了这种反应,而大肠被认为是大多数是厌氧的。总的来说,这些结果表明肠道微生物群具有从BBN生成BCPN的代谢能力。
为了鉴定能将BBN转化为BCPN的肠道细菌种类,我们在有氧和微氧条件下组装了一组来自小鼠肠道的564种无菌细菌培养物,并在相应的生长条件下测试了它们对BBN的氧化能力。利用活性分离物的全长16S rRNA基因的DNA测序分析,我们鉴定出属于8个不同细菌属(即大肠杆菌、假海螺菌、乳酸杆菌、葡萄球菌、枯草杆菌、小球菌、节杆菌和微球菌)和三个不同门(厚壁菌门、变形菌门和放线菌门;补充表21)的12个种。为了筛选在小鼠微生物组中以可检测水平存在的物种,并考虑到由于(微)好氧富集培养可能导致的潜在分离偏差,我们对盲肠、结肠和直肠样本进行了16S rRNA测序。然后,我们将样本中鉴定的扩增子序列变异体(ASVs)的参考序列映射到BBN转化细菌分离物的全长16S rRNA序列。这鉴定出属于四个不同属(大肠杆菌、乳酸杆菌、枯草杆菌和葡萄球菌;图3b)的八个ASVs。这种方法进一步使我们能够估计这些假定的BBN代谢细菌在肠道中的集体丰度,我们发现这些丰度很低(盲肠中0.48%,结肠中1.66%,直肠内容物中0.50%;图3c和补充表22)。这些观察结果表明,BBN的氧化是由肠道微生物群的一个独特亚群执行的,考虑到反应在大多数厌氧环境中对氧气的依赖性,这是意料之中的。
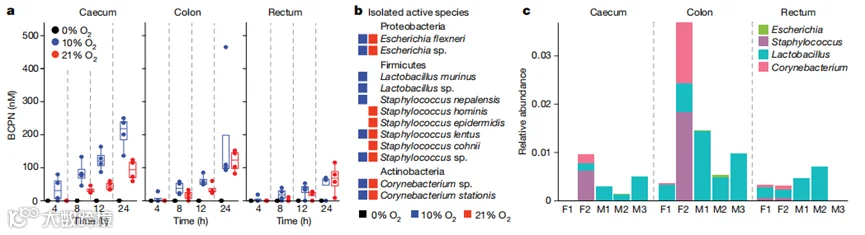
图3 | 来自小鼠的肠道细菌群落和分离物对BBN至BCPN的体外转化
人体肠道微生物群将BBN转化为BCPN
接下来,我们研究了人体肠道微生物群是否也能将BBN氧化为BCPN,尽管与小鼠肠道微生物群相比,其组成通常存在很大的差异性。为此,我们收集了11位志愿者的可培养粪便样本,并进行了16S rRNA测序分析。正如预期的那样,这些肠道微生物群与小鼠群落的确切分类学只有10.56%相同,确切的ASVs只有3.36%相同,只有一个可能活跃的细菌属(大肠杆菌)重叠(补充表25)。为了直接评估人体肠道微生物群的BBN转化能力,我们在厌氧、微氧和好氧条件下体外培养了这些粪便群落,与BBN共孵化,并在24小时后如上所述定量BCPN的产生。尽管我们通过菌落形成单位计数评估了群落的可比培养密度(扩展数据图4a和补充表24),但在十个群落中(由于技术原因排除了一个群落),只有六个在24小时BBN孵化后产生了BCPN,其中在微氧条件下活性最强(图4a和补充表23)。此外,这六个活跃的微生物群落显示出BBN氧化活性的显著差异,表明人与人之间的微生物群BBN代谢存在显著差异。为了解释这些个体间差异,我们探索了个体之间微生物组组成的总体差异,但没有发现基于粪便群落的BBN转化能力的任何分组。总的来说,这些结果表明,与小鼠肠道微生物群相似,尽管组成上存在明显差异,某些人体肠道微生物群也能够将BBN氧化为BCPN。
为了进一步确认人体微生物群在体内将BBN转化为BCPN的能力,我们在进行上述相同BBN暴露实验之前,用参与者G11的粪便微生物群对无菌小鼠进行了定植,这些“人类化”的小鼠(图1a)。肠道中BBN和BCPN的定量再现了在常规小鼠中观察到的代谢表型,也证明了人体肠道微生物产生的BCPN(图4b)。
为了鉴定将BBN转化为BCPN的人体肠道细菌,我们首先测试了16种不同的肠道细菌模式菌株,这些菌株涵盖了肠道中四个最丰富的门,在0%、10%和21%氧气的不同生长条件下,它们将BBN转化为BCPN的体外能力。在这些测试的菌株中,只有大肠杆菌ED1a能够产生BCPN(图4c),这证实了我们在人体微生物组中也发现了小鼠微生物组中唯一将BBN转化的属——大肠杆菌。为了鉴定不包括在模式菌株小组中的其他代谢BBN的人体肠道细菌,我们组装了来自不同粪便群落的个性化培养集合,总共有430种在微氧和好氧条件下分离的无菌细菌培养物。然后,我们测试了每种分离物氧化BBN的能力,结果从每个粪便群落中鉴定出最多15种转化BBN的物种(补充表33)。总的来说,我们从2个不同的门(厚壁菌门和变形菌门;扩展数据图4g)中分离出了25种独特的转化BBN的物种和18个不同的细菌属。值得注意的是,可以从我们社区BBN转化测定中整体不活跃的粪便群落中分离出几种转化BBN的物种。这可能由于这些转化BBN的细菌菌株在群落中普遍丰度较低,由于(微)好氧菌之间的竞争,在密集的群落环境中氧气供应有限,以及无菌和群落培养物之间不同的细菌生理学(即基因表达)所解释。为了鉴定在原始人类粪便样本中可检测到水平的细菌分离物,并考虑到由于(微)好氧富集培养可能导致的潜在培养偏差,我们将分离物的全长16S rRNA序列映射到16S rRNA微生物组数据,如上所述。我们能够映射出粪便微生物组的七种ASVs,属于三个不同的属(大肠杆菌、肠球菌和嗜血杆菌;扩展数据图4h和补充表33)。与小鼠微生物组一样,这些ASVs(假定的BBN代谢物)的相对丰度在不同群落中范围在0%到10.52%之间,平均为3.94%(扩展数据图4i)。然而,尽管假定的转化BBN的ASVs的相对丰度存在很大差异,我们没有发现这些ASVs的总相对丰度与粪便群落的BBN转化能力之间有任何相关性(扩展数据图4j)。这可能是因为群落环境对BBN氧化很重要,或者是由于16S rRNA扩增子测序的分辨率有限,可能无法完全反映物种和菌株水平的BBN代谢差异。尽管这些细菌的相对丰度无法预测微生物群落氧化BBN的代谢潜力,但我们的数据表明,人体肠道微生物群BBN氧化能力存在显著的个体间差异,并且可以分离出具有代谢BBN能力的不同的(微)好氧人体肠道微生物。
我们可以从小鼠和不同个体(图3b和4d)中分离出大肠杆菌的不同菌株,这一事实表明将BBN转化为BCPN的能力在这一细菌属的不同菌株中是保守的。为了验证这一假设,我们对来自不同个体的七株大肠杆菌的基因组进行了测序,证明它们确实是不同的菌株(扩展数据图4k和补充表34及35)。此外,我们测试了之前分离的95种不同的大肠杆菌菌株,它们将BBN转化为BCPN的能力(图4e和补充表36)。我们发现所有测试的菌株都能够代谢BBN,这表明氧化BBN的能力在大肠杆菌菌株中是共享的,并且是不同大肠杆菌种类所共有的。为了测试代表性人体大肠杆菌分离株在体内的BBN代谢,我们将无菌动物与我们的一株分离株(大肠杆菌flexneri)进行单菌落定植,比较这些单菌落定植和无菌动物在饮水中给予BBN 10天后的肠道BBN代谢。gBBN和BCPN的测量再次证实了我们的发现,即肠道细菌在常规和“人类化”小鼠的肠道中代谢BBN,导致BCPN在下肠道积累和吸收(图4f,g和补充表37及38)。此外,为了更好地模拟肠道内的物种间相互作用,我们还用两种不代谢BBN的细菌模式菌株Coprococcus comes和Flavonifractor plautii(图4c)以及添加了分离的BBN代谢大肠杆菌flexneri菌株的三菌株群对无菌小鼠进行了定植。在宿主代谢没有差异(图4h)的情况下,三菌株群导致下肠道BCPN积累,与缺乏BBN代谢细菌的两菌株群形成对比(图4i,j和补充表39及40)。总的来说,这些结果表明特定的人体肠道细菌能够在肠道中代谢BBN,这导致了亚硝胺致癌物的毒代动力学改变。
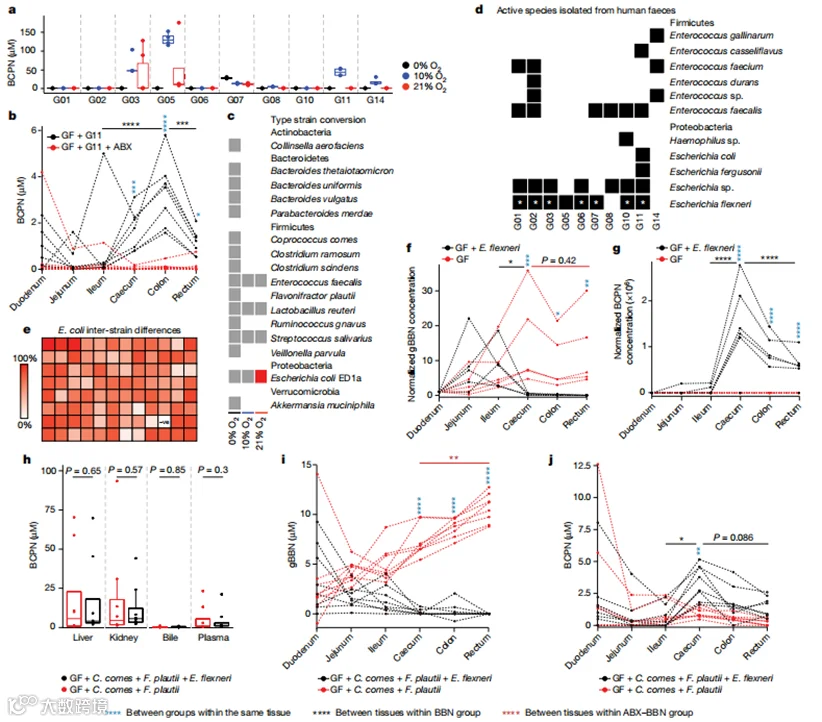
图4 | 个体间在BBN至BCPN转化的差异以及单一分离物在体外和无菌小鼠模型中的代谢活性。
微生物群影响其他亚硝胺
在确定BBN的毒代动力学被肠道微生物代谢改变后,我们质疑其他相关的亚硝胺致癌物是否也会受到肠道微生物群的类似影响。为此,我们用三种额外的亚硝胺致癌物(N-乙基-N-(4-羟基丁基)亚硝胺(EHBN)、N-亚硝基二丁胺(DBN)和N-丙基-N-丁基亚硝胺(PBN))对常规和经ABX处理的动物进行了24小时的代谢笼实验(图5a)。正如我们用BBN和BCPN的发现所预期的,我们发现微生物群也改变了这些额外测试的亚硝胺致癌物的毒代动力学,导致通过ABX处理减少肠道细菌负荷后,亚硝胺及其氧化代谢物的尿液分泌减少(图5b-d和补充表41)。
由于EHBN通常用于研究小鼠模型中化学诱导的膀胱肿瘤发生,我们调查了微生物群对毒代动力学的观察影响是否也会在功能上转化为改变的膀胱肿瘤发生。为此,我们进行了为期20周的EHBN长期暴露实验。与我们对BBN的发现一致,我们的观察还揭示了EHBN暴露的小鼠在完整的微生物群中发生了膀胱致癌,而在接受ABX-EHBN处理的动物中我们观察到肿瘤发生减少(图5e)。
此外,我们观察到EHBN的氧化代谢物N-乙基-N-(3-羧丙基)亚硝胺(ECPN)在非ABX处理的小鼠中只在大肠中积累,与BBN暴露后BCPN的积累类似(图5f)。最后,对ABX-EHBN处理的小鼠中葡萄糖醛酸化EHBN的测量以及在EHBN处理的小鼠中以ECPN形式的总EHBN的质量平衡表明,观察到的EHBN毒代动力学的改变可能是肠道细菌脱结合和氧化反应的结合(图5g,h)。总的来说,这些结果表明肠道微生物代谢可以改变亚硝胺致癌物的毒代动力学,并影响化学诱导的肿瘤发生。
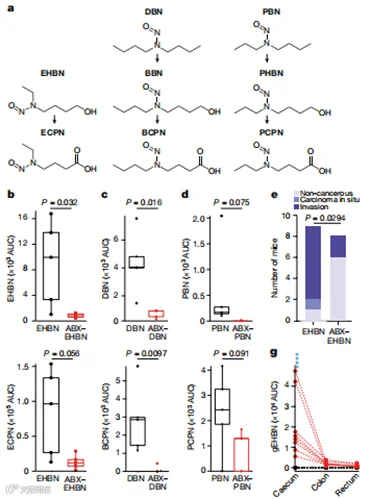
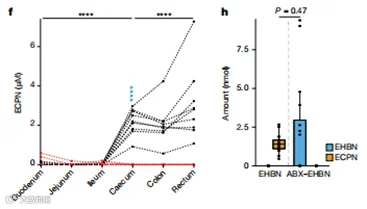
图5 | 微生物群依赖的与BBN相关的亚硝胺致癌物的毒代动力学
讨 论
在这项研究中,我们证明了微生物对环境致癌物的生物转化是癌症发展的一个促成因素。我们在机制上展示了肠道微生物群如何通过微生物对环境致癌物的代谢,促进化学诱导的肿瘤发展并在远端器官加速癌症进展。通过将活跃的人类粪便群落以及单菌株分离物和有无活跃BBN转化菌株的合成细菌群落接种到无菌小鼠中,我们可以像在常规小鼠中一样再现微生物将BBN及其衍生物转化为BCPN的毒代动力学。我们证明了细菌代谢的作用不仅在于通过脱糖基化再生亲本致癌物,而且还通过一个两步机制积极产生致癌代谢物。我们发现个体微生物群代谢致癌物的能力存在显著差异,这表明存在人与人之间微生物群对化学诱导肿瘤发展贡献的差异。然而,微生物组中BBN代谢物种的丰度无法解释相应微生物群落的致癌物代谢活性。这可能是由于群落环境可能决定个别细菌菌株的致癌物代谢活动,并建议进一步研究以更好地理解微生物群对环境污染物的代谢。
综合起来,这些结果补充了之前涉及肠道微生物组在癌症发展中的研究,并提供了一个方法论和概念框架来研究微生物组致癌物代谢及其对毒代动力学和肿瘤发展的影响。未来的研究将基于这些发现,旨在鉴定微生物组编码的化学诱导癌症发展的易感风险因素,并实现针对操纵个体微生物群以预防癌症的合理策略。
往期推荐
