
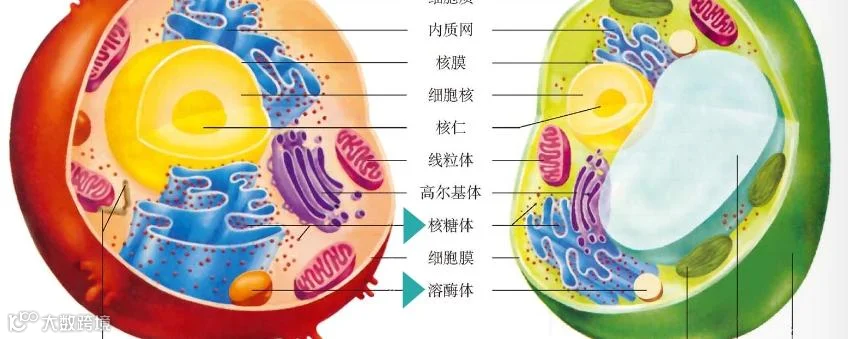
Fig. 4 线粒体质量控制途径
如果把线粒体放在正确的位置似乎是一项艰巨的工作,那么神经元在其深远的末端维持健康线粒体池的能力是无与伦比的。线粒体DNA缺乏对甲基化的保护,并且暴露于高水平的ROS。Ca2+和其他离子、神经递质、压力源和毒素由于神经元活动、兴奋性毒性和衰老而积聚在突触内,使突触线粒体容易受损。神经元的高能量需求进一步加剧了线粒体的脆弱性。受损的线粒体不仅在突触局部有害,还可能引发全身免疫反应。
突触线粒体如何在稳定状态和压力下保持健康,仍然是未来研究的重要领域。有几个关键问题特别令人感兴趣;例如,哪种类型的线粒体质量控制是在突触局部实施的,它是否受到突触活动和可塑性的调节?此外,远端突触距离产生新蛋白质的细胞体很远,质量控制的分子参与者是如何快速传递到远端突触中受损的线粒体的?
一些突触的偏远位置表明,局部蛋白质翻译对质量控制至关重要。信使核糖核酸可能已经被输送并储存在远端突触中,一旦迫切需要这些蛋白质,就可以开始翻译。有人提出,某些核编码的线粒体蛋白的mRNA附着在OMM上。快速的局部翻译可以提供新产生的线粒体蛋白,使年老受损的线粒体恢复活力,并维持线粒体的功能。局部蛋白质供应也可以充分维持线粒体表面的泛素-蛋白酶体系统,以去除有缺陷的OMM蛋白质或在进口过程中停滞的蛋白质,或快速提供线粒体蛋白酶和伴侣蛋白,以清除线粒体内错误折叠或变性的蛋白质(Fig. 4a)。局部蛋白质翻译可以进一步维持调节选择性去除线粒体缺陷部分的关键蛋白质参与者,包括线粒体类核富集自噬体(autophagosome,APs)、线粒体衍生的囊泡和隔室(mitochondria-derived vesicles and compartments,MDVs and MDCs)以及OMM阳性结构(structures positive for OMM,SPOTs)的产生(Fig. 4b),或者可以控制线粒体自噬,整个线粒体通过线粒体自噬被引导到溶酶体进行降解(Fig. 4c)。
线粒体自噬是一种线粒体选择性自噬,可以发生在体外培养的和在体的神经元中。在线粒体自噬过程中,使用了与其他细胞器相同的自噬机制来启动、扩张和吞噬受损的线粒体,当然还需要额外的分子参与者(Fig. 4c)。研究的最好的线粒体自噬途径是由PINK1和Parkin介导的。线粒体去极化阻断Ser/Thr激酶PINK1的线粒体输入,使其在OMM上稳定。PINK1随后磷酸化多个OMM蛋白和相邻的泛素分子,导致胞质E3连接酶Parkin活化并募集到线粒体表面。Parkin被PINK1的磷酸化进一步激活,并继续泛素化更多的OMM蛋白,使通路升级。这种前馈机制在线粒体表面引起广泛的磷酸化和泛素化事件,吸引自噬机制(Fig. 4c)。线粒体也可以通过PINK1–Parkin非依赖性线粒体自噬途径以及非选择性大自噬清除。
此外,与大多数其他类型的细胞一样,线粒体可能在突触处经历动态裂变和融合,以丢弃、刷新或交换其内容物(Fig. 4d)。分裂和融合是由线粒体膜蛋白介导的:动力蛋白相关蛋白1(dynamin-related protein 1,Drp1)及其用于分裂的受体,以及用于融合的丝裂融合蛋白(mitofusin,MFN1和MFN2)和视神经萎缩1(optic atrophy 1,OPA1)(见最近对分裂和融合机制的综述)。
除了局部蛋白质供应外,基于微管的顺行转运可能从细胞体递送更多的溶酶体、自噬体或囊泡中包装的调节蛋白,以支持线粒体质量控制途径。受损的线粒体也可能被突触末端的自噬体局部吞噬,然后运回胞体,或者只是从突触喷出到相邻的非神经元清除细胞进行降解。
监测机制的多样性非常适合突触线粒体的可塑性,并开启了一场对话,以解决如何在完整的神经元回路中的独特突触中产生不同类型的线粒体质量控制的基本问题。例如,一些MDVs出现在轻度氧化应激和先前的线粒体自噬下,这是由长期和过度的损伤触发的。如果这两种途径都用于远端神经炎,可能会有分子传感器来切换一个“打开”和另一个“关闭”。
最近的一项研究表明,某些类型的MDVs的生物发生取决于线粒体膜最初微管依赖性突起的Miro(Fig. 4b)。相反,去极化触发的线粒体自噬需要Miro的蛋白酶体降解,以使线粒体与微管解偶联(Fig. 4c)。尽管Miro可以作为在这两种质量控制途径之间切换的分子按钮,但其调节机制仍有待解决。
神经元可能需要在突触线粒体受损时迅速做出决定,考虑到能量消耗和代谢需求的所有因素,同时努力保持其电活动。多学科方法,包括可扩展的信使核糖核酸生物传感器、新合成的蛋白质、活性氧、代谢产物、ATP和MDV、MDC、SPOT和线粒体自噬途径的敏感报告子,结合超分辨率显微镜、双光子解锁的单个突触刺激和常规电生理技术,将提供一系列优秀的工具,在体内和培养的神经元中以单一突触分辨率解决这些问题。
靶向突触线粒体的治疗潜力
维持突触能量蓝图的失败对神经元是有害的。单个突触的能量短缺、Ca2+和ROS的积累可能导致突触损失。突触结构和功能的进行性丧失是轴突收缩和细胞死亡之前的早期迹象,也是许多神经退行性疾病的共同特征,如阿尔茨海默病(Alzheimer’s disease,AD)、肌萎缩侧索硬化症(amyotrophic lateral sclerosis,ALS)和帕金森病(Parkinson’s disease,PD)。
人类遗传学研究发现,编码线粒体膜蛋白的基因存在变异,这些基因介导各种神经系统疾病中的线粒体运输、动力学和适应,如脑病(DRP1和TRAK1)、2A型Charcot-Marie–Tooth病(MFN2)、帕金森病(OPA1和MIRO)和常染色体显性视神经萎缩(OPA1)。PINK1和PARKIN的突变,其产物在线性轴上起作用,以控制MDV和线粒体自噬途径(Fig. 4b,c),导致早发性隐性PD。此外,其产物介导线粒体自噬的视神经磷酸酶(optineurin,OPTN)和TBK1的变体(Fig. 4c)与ALS相关。强有力的遗传证据不仅表明,未能维持线粒体是神经病理学的直接原因,而不是其他神经元功能障碍的结果,而且还表明,靶向这些蛋白质和途径可能对疾病干预有效。

许多神经退行性疾病涉及年龄依赖性神经元损失。随着蛋白质组、脂质体和代谢组的变化,线粒体功能在大脑衰老过程中下降。即使在症状出现之前,识别晚发神经退行性变的早期指标或预测因素,对于早期干预以提高治疗效果尤其有价值。突触线粒体行为的变化很可能是神经元灾难的最初迹象之一,并可能在分子水平上得到反映。神经元的分子缺陷可能在外周组织中是保守的,这可能是生物标志物开发的极好候选者。
最近的研究表明,Miro1对PD模型中线粒体去极化诱导的降解具有耐药性,并且这种表型在皮肤成纤维细胞、诱导多能干细胞(induced pluripotent stem cells,iPSC)和来自PD患者的神经元中保留。重要的是,尽管在大型队列研究中,Miro1缺陷在统计学上与帕金森病患者和无症状遗传携带者显著相关,但它并不是发生在每个患有该疾病或遗传携带者身上。Miro1的长期滞留会导致受损线粒体的阻滞和清除延迟,进而导致氧化应激,最终导致PD神经元的细胞死亡。继续努力在临床可得到的组织(血液或皮肤)中寻找可以标记疾病前驱期或早期的分子事件,将使患者分层更加准确,提高药物试验的成功率,并改变临床护理。
