
循环系统类器官——血管
血管是循环系统的组成部分,负责将血液输送到机体各处。血管将血细胞、营养物质和氧气输送到身体组织,同时带走组织中的废物和二氧化碳。血管是维持生命所必需的,身体的所有组织都依赖于它们的功能。
血管类器官的研究和应用发展可从大脑血管、骨髓血管、淋巴管及遗传驱动器官型等进行初步了解。
大脑血管
脑血管系统高度专业化,其中星形胶质细胞与血管紧密结合,并与内皮细胞结合形成血脑屏障。小胶质细胞作为专门的免疫细胞,有助于神经发生并调节损伤的恢复。
2013年,Lancaster等人[1]发布了第一个脑类器官方案,脑类器官的一个关键方面是引入脉管系统来模拟血脑屏障,并提供更好的营养支持,从而促进较大类器官的生长和成熟。研究者在悬滴培养板中同时接种原代脑ECs、原代周细胞和原代星形胶质细胞,发现ECs是调节其他细胞类型增殖的重要组成部分。Sun等人使用血管类器官和脑类器官的共培养获得了血脑屏障样特征。通过Wnt信号向中胚层胚状体的初始分化,随后使用BMP4、bFGF进行血管诱导,然后嵌入基质胶中获得血管类器官。在2个血管祖细胞类器官之间神经上皮胚状体共包埋后,这些梭形细胞在神经血管成熟培养基中进一步培养。延长培养获得了血脑屏障特征。
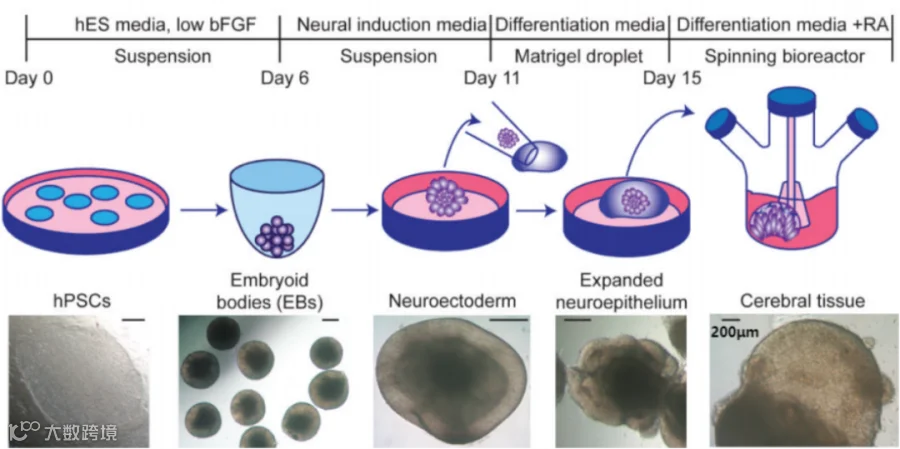
大脑类器官培养流程格式件节点明场图[1]
骨髓血管系统
骨髓是成年人造血干细胞的主要来源[2]。Giger等人[3]使用基于微孔的方法,将ECs和间充质干细胞共同接种以形成球状体。发现双细胞类型器官中自我更新的间充质干细胞数量较多。ECs自组织成网络。来自这些球状体的间充质干细胞可以分化成脂肪细胞、软骨细胞和前成骨细胞。在形成3天的类器官上接种造血干细胞后,细胞归巢并迁移到类器官内部,模拟了体内数据,即注射到受辐照的小鼠宿主中的造血干细胞将归巢并重建骨髓。
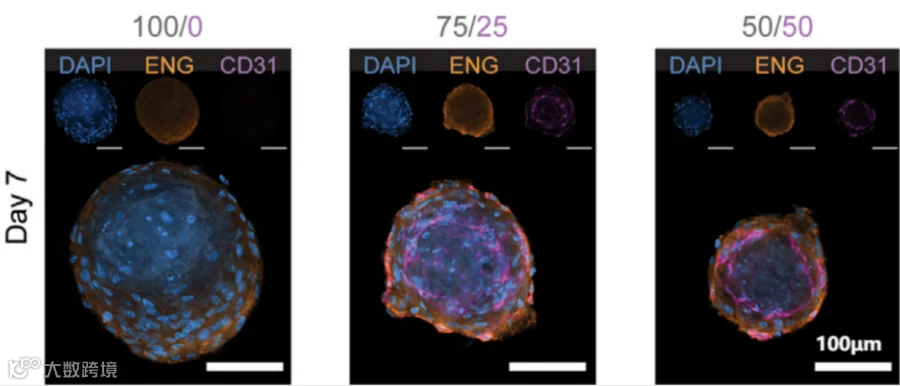
间质细胞与内皮细胞不同比例共培养7天后免疫荧光染色[3]
在另一种方法中,诱导多能干细胞最初聚集并诱导为中胚层聚集体[4]。通过添加BMP4、FGF2、VEGF-A、SCF和Flt3,类器官被包埋在胶原蛋白/基质胶混合物的水凝胶中。这些类器官显示出分化为腔化血管以及造血干细胞与血管的结合,类似于骨髓。单细胞测序分析证实了几种造血细胞和基质细胞亚型的存在[5]。轨迹分析表明,分化的细胞起源于体内也存在的骨髓分化的主要途径。
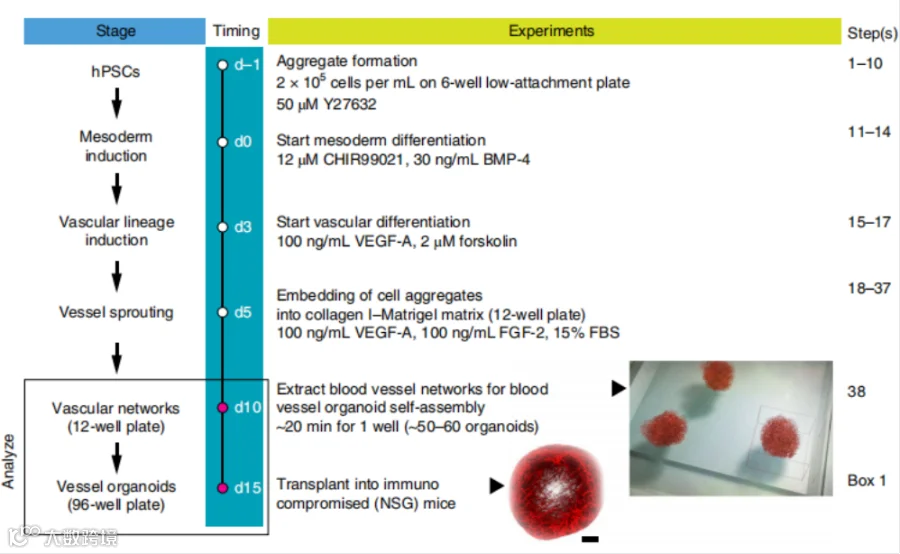
血管类器官培养流程[4]
淋巴管
淋巴管是微血管的一个特殊亚型[6]。由于淋巴管在免疫学、组织引流和疾病过程中起着重要的作用,众多研究者已经在淋巴管背后的基本生物学方面付出了很多努力,目前的体外方法包括在不同流量和ECM底物下的细胞形态评估[7],屏障功能测试[8],运输测试[9],渗透性测试等[10]。
癌细胞已经被证明可以分泌前淋巴管生成因子,淋巴管的形成是由淋巴管内皮细胞上VEGFR-3的表达调节的,并由配体VEGF- c /-D激活,淋巴管生成模型依赖于淋巴细胞的利用[11]。目前还没有能够准确地概括淋巴血管生成和淋巴管生成的体外模型,这种模型的一个主要障碍是淋巴ECs的起源。在胚胎发育过程中,淋巴EC源自Prox1+/Lyve1+静脉ECs群[12]。静脉血管的要求引起一个问题,即目前可用的BVO方案是否可以进行修改以完成体外淋巴血管生成。
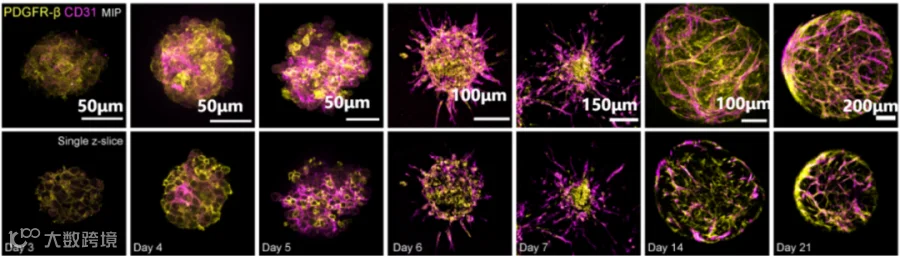
血管生成过程免疫荧光[12]
遗传驱动器官型
在Dailamy A等人关于大脑血管类器官的研究中[13],研究人员通过过表达NEUROD1以诱导神经血管生成,同时过表达MYOD1B+AF60C来创建肌血管类器官。在NEUROD1诱导第15天的BVOs中,检测到表达MAP2+的神经元,在类器官内聚集分布。相同条件下的肌血管类器官中也观察到类似的情况。表明在长时间培养后,神经元标记物 MAP2、VGLUT2、BRN2和 FOXG1上调。神经血管类器官的进一步表征显示神经元能够自发放电,证明了神经元组织的功能性。
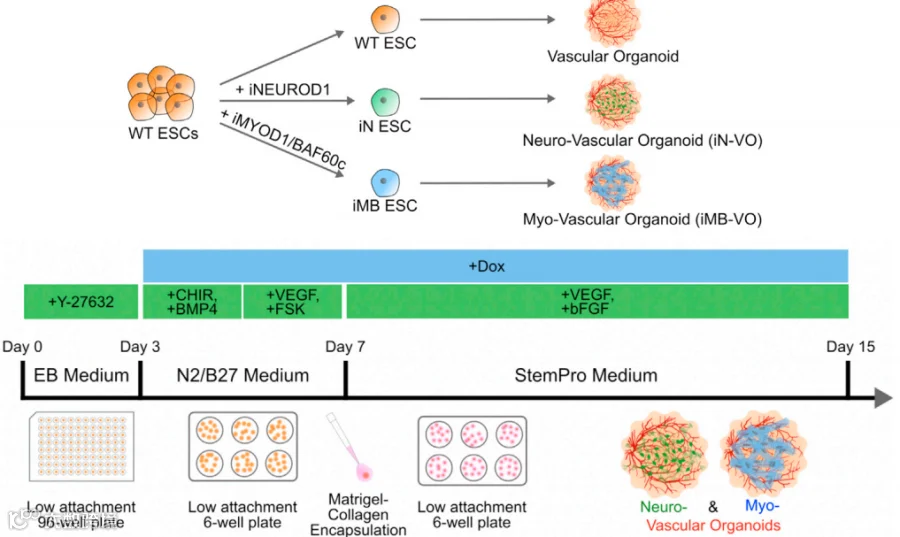
细胞处理及血管类器官培养流程示意图[13]
在众多与血管类器官联合生长应用的类器官研究中,有很多疑问和困难需要解决,如本文述的淋巴管类器官的研究,以及血管形成的能力和器官型血管系统形成方案的确立等。因此,血管类器官及组织联合类器官的研究仍在继续。
公司简介


微信号|一米生物
服务热线|400-097-3606
