
三羧酸循环的中间产物α-酮戊二酸(AKG)参与细胞内的多效性代谢和调节途径,包括能量产生、某些氨基酸的生物合成、胶原生物合成、基因表达的表观遗传调节、氧化还原稳态的调节,以及有害物质的解毒。α-酮戊二酸还是AKG依赖性双加氧酶的必需底物,其家族包括DNA和组蛋白去甲基化的主要酶,如DNA的去甲基酶TETS和含有Jumonji C结构域的组蛋白赖氨酸脱甲基酶,如KDM2-7。TET1-3催化AKG氧化脱羧酶,将5-甲基胞嘧啶转化为5-羟基甲基胞嘧啶,从而触发DNA中去甲基化GpC位点[1]。jmcc-结构域组蛋白去甲基化酶可从组蛋白的明显甲基化赖氨酸残基中去除甲基,后者是一个关键的“表观遗传标记”,它参与转录活性染色质或非活性染色质的形成[2]。
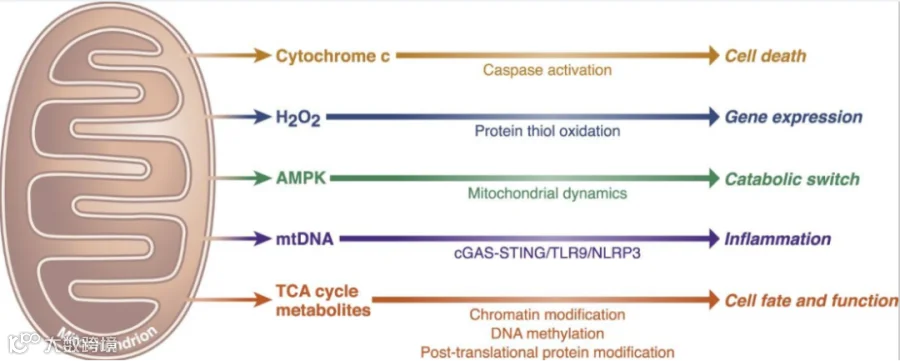
线粒体的重要信号传导功能[2]
维生素C是一种人体必需的营养物质,是重要的抗氧化剂,并且在体内参与了胶原蛋白、儿茶酚胺类及肉毒碱等物质的合成。更重要的是维生素C是依赖金属离子以及酮戊二酸的双加氧酶类(dioxygenases)的辅酶,如脯氨酸羟化酶(prolyl-4-hydroxylase)会在缺乏维生素C的情况下造成胶原合成和成熟受阻,导致坏血病[3]。它负责电子传递,可以及时将反应生成的Fe4+还原形成Fe2+,而其自身被氧化形成脱氢抗坏血酸(dehydroascorbic acid, DHA)。而在表观遗传中,维生素C作为辅酶的就是DNA的去甲基酶TETs和含有Jumonji C结构域的组蛋白赖氨酸脱甲基酶[4]。
裴端卿团队发现组蛋白去甲基酶Jhdm1a/1b是维生素C下游体细胞重编程的关键效应因子,通过抑制Ink4/Arf位点,在重编程过程中加速细胞周期进程并抑制细胞衰老。由于间充质干细胞向上皮细胞转化(MET)步骤受阻,Tet缺乏的小鼠胚胎成纤维细胞不能被重新编程。缺乏DNA糖基化酶(TDG)的小鼠胚胎成纤维细胞的重编程也同样受阻。他们发现只有3个Tet基因都敲除的小鼠胚胎成纤维细胞才不能重编程,任意一个Tet基因未敲除都不能达到这个效果,而这一效果是对于MET至关重要的miRNA(如miR-20c)的表观遗传活化有障碍造成的[5]。他们发现3种TET都被敲掉的小鼠会发生与轴性中胚层成熟受损相关的原始条纹缺陷和旁轴中胚层缺陷,这是Nodal抑制剂Lefy1和Lefy2基因的DNA甲基化的升高导致表达减少,导致过度活跃的Nodal信号产生的,当Dnmt3a和Dnmt3b受到阻碍时,Lefy-Nodal信号传导以及缺陷小鼠的特定表型都会得到恢复[6]。
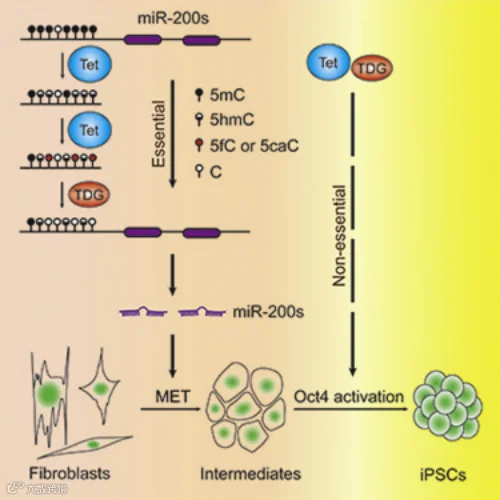
Tet 和 TDG 介导体细胞重编程中EMT所必需的 DNA 去甲基化[5]
原肠胚形成是发育的一个基本的早期步骤。作用于基因调控元件,TET介导的5-甲基胞嘧啶氧化平衡从头甲基化,以确保其对上游信号通路驱动的转录激活的反应,在缺乏TET的情况下,异常甲基化会导致Lefy-Nodal回路失调。
之后他们发现绿藻衣藻(reinhardtii)中有一个5-甲基胞嘧啶修饰酶(CMD1),它是TETs的同系物,不同的是,TETs以2-酮戊二酸为辅酶,CMD1以维生素C为辅酶[7]。接着他们继续研究CMD1催化的DNA修饰新方法—C5-甘油基甲基胞嘧啶(5gmC),CMD1体外DNA结合实验表明CMD1对不同长度、结构和5-甲基胞嘧啶水平的DNA具有可比的结合亲和力,并且对含有5mCpG的DNA表现出比含有5mcCP-、5mCpA-和5mCpT-的DNA适中的底物偏好。
维生素C的内酯形式与活性位点结合,并以不同于Fe2+/2-OG依赖性双加氧酶中2-OG的方式单配位Fe2+。参与Fe2+和维生素C结合的关键残基的突变使酶活性丧失,参与DNA结合的关键残基的突变使酶活性显著降低,揭示了CMD1如何识别DNA底物和利用维生素C作为共底物的分子基础[8-9]。
参考文献
公司简介


微信号|一米生物
服务热线|400-097-3606
