
中枢神经系统是脊椎动物体内最复杂的系统之一。胚胎发育过程中,中枢神经系统的发育受到复杂而又次序严谨的调控。为了研究早期胚胎中多能干细胞的适当的神经启动,利用全基因组芯片测序和RNA测序分析,景乃禾院士团队证明转录因子Pou3f1是神经促进基因的上游激活因子,并且能够抑制神经抑制信号[1]。Pou3f1可以直接结合SOX2等神经谱系基因和BMP、Wnt等神经抑制信号下游靶点,是神经命运决定过程中内在转录因子和外在细胞信号的关键双重调节因子。
同济大学孙方霖团队研究了Noch信号对果蝇的正常发育与肠道稳态的重要作用[2]。他们发现异染色质蛋白1c(heterochromatin protein 1c, HP1c)是果蝇肠道内稳态的重要表观遗传调节因子,HP1c通过直接与Notch信号传导的关键转录因子无毛抑制因子[Su(H)]相互作用抑制Notch靶基因的转录。在果蝇中表达人HP1可以挽救HPIc缺失引起的表型γ,表明HPIγ在果蝇中类似于HPlc的功能。
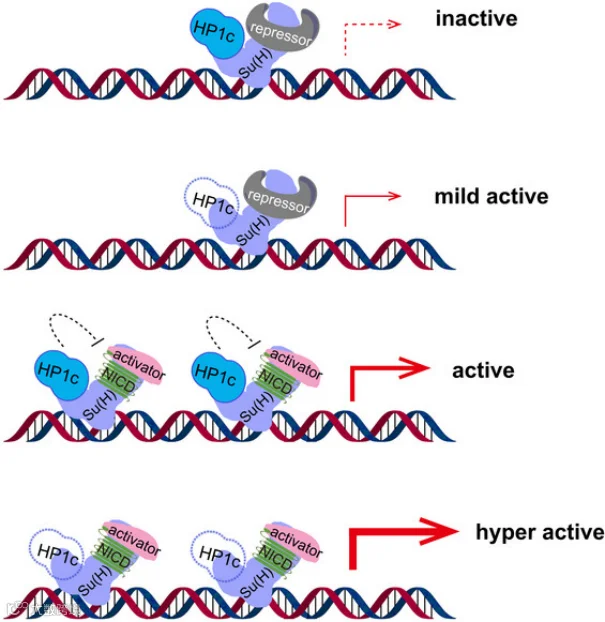
HP1c 调控 Notch 信号通路的模型[2]
染色质状态和细胞代谢都与细胞命运决定相关,但它们之间有什么相互作用呢?中国科学院上海营养与健康研究所的金颖团队通过全基因组siRNA筛选确定PHB是人胚胎干细胞自我更新的一个重要因子[3]。PHB与HIRA、组蛋白H3.3伴侣形成蛋白复合物,并稳定HIRA复合物的蛋白质水平。PHB和HIRA共同作用控制人胚胎干细胞中组蛋白H3.3的沉积和基因表达。PHB和HIRA调节异柠酸脱氢酶基因启动子处的染色质结构,从而促进转录,促进其产生胚胎干细胞(Embryonic stem cell,ESCs,简称ES、EK或ESC细胞)命运调控的关键代谢产物—α-酮戊二酸。
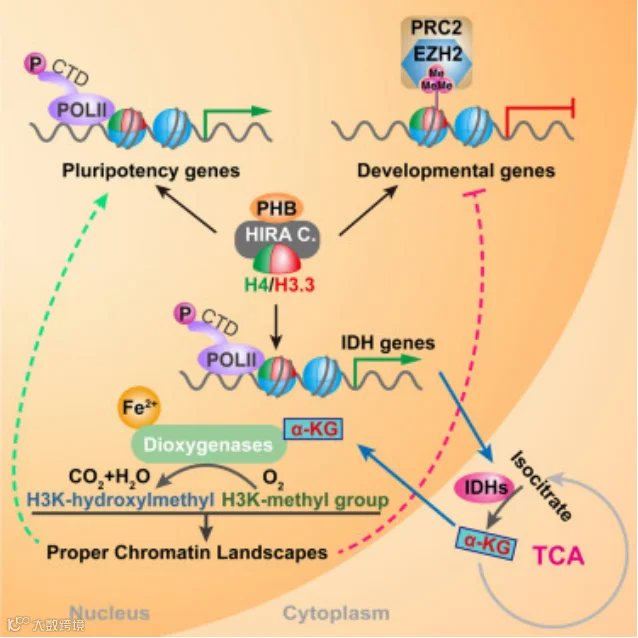
PHB 与 HIRA 复合物结合,调控人类 ESC 的表观遗传代谢[3]
c-Jun是一个著名的癌基因,但是它在胚胎中细胞命运决定中也发挥着关键的作用。通常认为癌基因在重编程过程中是一个促进因素,但裴端卿团队发现c-Jun实际上是重编程的一个重要障碍[4],他们的实验表明,与重要的多能性基因OCT4对应,c-Jun是一个体细胞状态的守护者,而OCT4是干细胞状态的守护者。抑制c-Jun可以促进体细胞重编程,而抑制OCT4则可以促进干细胞分化。他们的实验还表明c-Jun的显性阴性突变体和Jdp2都可以在重编程中替代OCT4,进一步证实了他们此前的理论。
除了基因的影响以外,代谢产物对细胞命运的影响也非常大,刘兴国团队用高覆盖率的脂质组学方法研究了脂质代谢在多能干细胞中的作用[5]。磷脂酰乙醇胺(PE)合成的CDP-Z醇胺(CDP-Etn)途径在重编程的早期阶段是必需的。CDP-Etn通路以Pebp1依赖性方式抑制NF-KB信号传导和间充质基因,从而加速间充质到上皮转化(MET)并增强重编程。PE与Pebp1的结合增强了Pebp1与IKK的相互作用α/β降低IKK的磷酸化α/β。CDP-Etn-Pebp1轴在肝细胞分化中与EMT/MET相关,提示Etn/PE是一种广谱的MET/EMT调节代谢产物。说明了磷脂在细胞命运中的重大作用[6]。
参考文献
公司简介


微信号|一米生物
服务热线|400-097-3606
