
越来越多的研究表明,对于肿瘤干细胞的发生与功能维持,表观遗传学的调控机制可能发挥着极其重要的作用。肿瘤的易感性、进展和异质性均与干细胞在肿瘤晚期阶段的表观遗传学紊乱有密切联系,表观遗传学紊乱同时也参与了成体干细胞向肿瘤干细胞的转变[1]。这些表观遗传调控可以影响肿瘤干细胞的自我更新和分化形成新肿瘤的能力。表观遗传调控癌变重编程、肿瘤干细胞的自我更新,以及针对肿瘤干细胞表观调控机制的靶向治疗等,已成为肿瘤生物学研究的重点研究方向。
另外,已经有许多研究表明肿瘤干细胞带有异常的表观遗传学修饰,这些修饰通过影响 Wnt/β-catenin、Hedgehog、Notch 和TGF-β/BMP 信号通路,调控肿瘤干细胞的自我更新和分化能力,进而改变肿瘤细胞生长、恶化、转移、复发及抗药的能力[2-5]。
1.表观遗传调控分类
表观遗传学调控主要包括 DNA 甲基化、组蛋白修饰、染色质重塑和非编码RNA(non-coding RNA, ncRNA)等。上述表观遗传学修饰类型既能独立发挥功能,又可以相互联系、共同作用,产生“协同效应”[6-8],通过调节关键基因的表达,维持稳态,而当其出现异常改变时。则会引起多种疾病,包括肿瘤。
2.DNA 甲基化
DNA 甲基化是最常见的一种表观遗传学调控方式,是肿瘤发生的起始事件,先于基因突变。DNA 甲基化大多发生在富含 CG 的基因区域,由 DNA 转移酶I(DNMTI)和 DNA 转移酶 3A、3B(DNMT3A、3B) 催化形成,通过与甲基化结合蛋白(methyl-binding protein,MBP)识别、结合,招募转录共抑制因子,进而介导转录抑制,是表观遗传学调控基因表达最常见的机制之一[9]。它在基因表达沉默、印记和X染色体失活中发挥着重要的作用,对于肿瘤的发生发展,肿瘤干细胞的调控都起着重要的作用[10,11]。
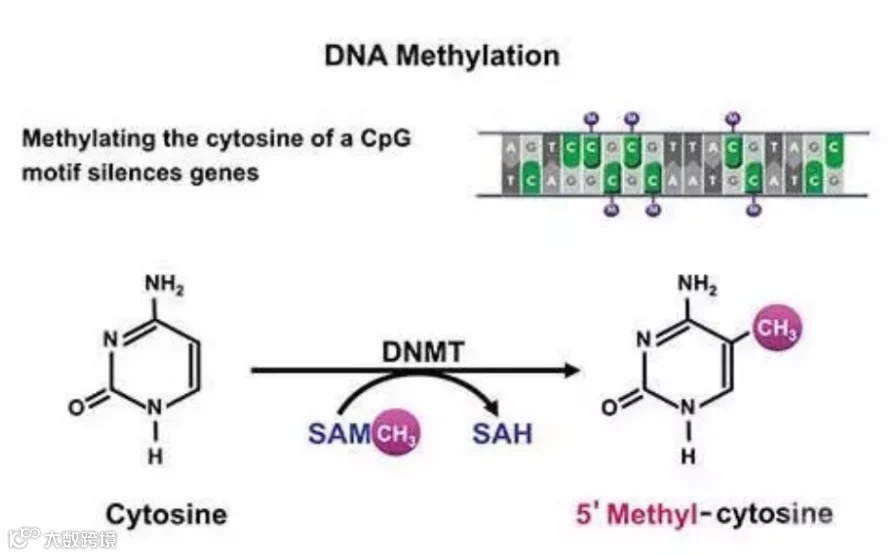
图1 DNA甲基化示意图
3.组蛋白修饰
组蛋白修饰常发生在组蛋白的氨基端,由于暴露在染色质外面,可接受各种化学基团的修饰。组蛋白修饰又称组蛋白密码,决定着基因的开放与否,目前研究最广的是组蛋白 H3、H4 上赖氨酸 K 的乙酰化和甲基化。当组蛋白高乙酰化时,基因转录激活;而甲基化则根据其甲基化位点的不同有所不同。一般认为,H3K9(组蛋白 H3 第 9 位赖氨酸残基)和 H3K27 甲基化与基因转录抑制相关,而 H3K4、H3K36 和 H3K79 与基因转录激活相关[12]。
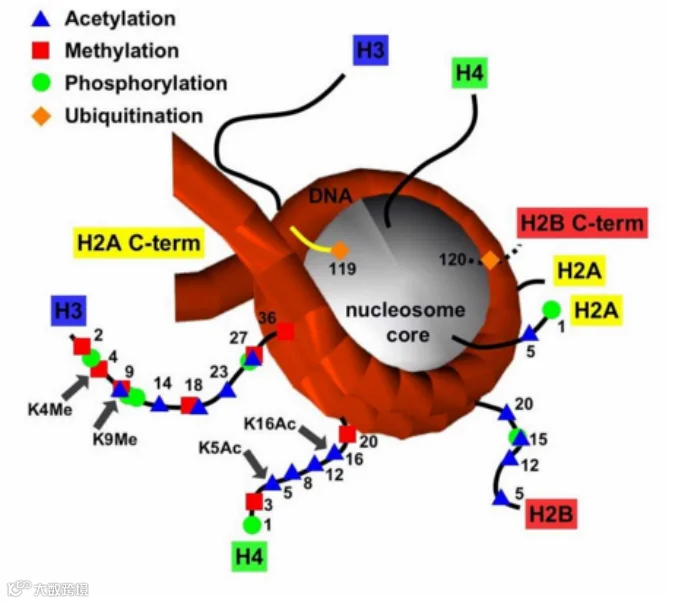
图2 组蛋白修饰示意图
4.染色质重塑
染色质重塑是另一种重要的表观遗传学调控机制,主要涉及4种 ATP 依赖染色质重塑因子 SWI/SNF、CDH、INO80、ISWI。重塑因子间相互作用,通过形成重塑复合物,如 BAF(SWI/SNF)、NURD、ISWI、CDH1 及 Tip60-p400 等,改变染色质的缠绕密度从而影响转录因子与 DNA 序列的结合,进一步调控基因表达。染色体结构的变异已经被报道与胚胎干细胞种类和分化能力的调节有关,其对于肿瘤干细胞的调控也可能会起作用[13]。
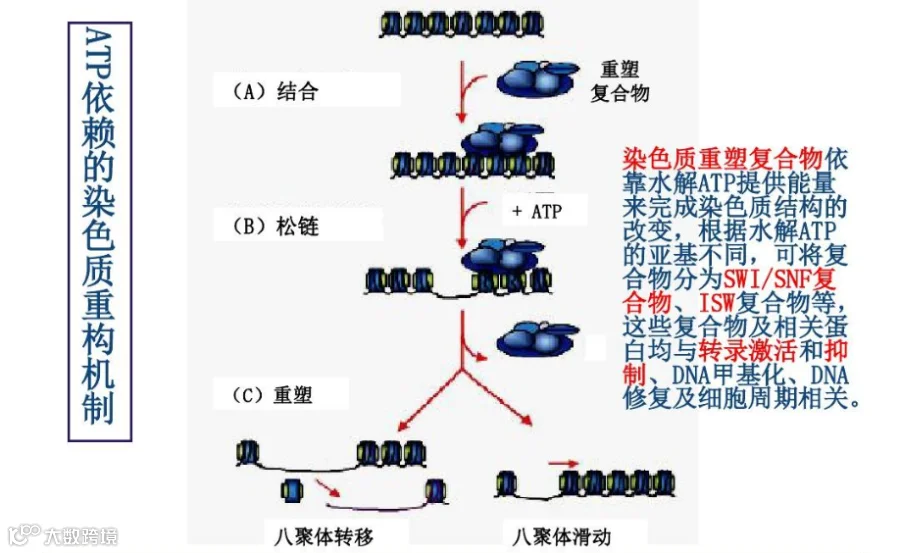
图3 染色质重塑示意图
5.ncRNA 和 miRNA
非编码 RNA(nCRNA)也属于广义上的表观遗传学修饰范畴。ncRNA 在基因表达中起重要的作用,它们根据其大小可分为长链 ncRNA 和短链 ncRNA。长链 ncRNA 在基因簇以至于整个染色体水平中发挥着顺式调节作用,短链 RNA 可能扮演着保护基因组稳定性的角色,包括微小 RNA(microRNA, miRNA)在基因表达调控中发挥非常重要的作用。许多已知的致癌性 miRNA 在未分化的干细胞中表达,而在分化的组织中表达降低。肿瘤干细胞中表达减少的 miRNA 称为抑癌性 miRNA,可负性调节肿瘤干细胞的自我增殖和致瘤能力,抑制干性[14]。
6.其他表观遗传学机制
除了 DNA 甲基化、组蛋白乙酰化、染色质重塑和 ncRNA,肿瘤干细胞也受其他表观遗传学机制调节,例如基因印记缺失(loss of imprinting,LOI)。LOI 是指正常不表达的等位基因异常激活或正常表达的等位基因异常沉默。它是人类肿瘤中常见的遗传学改变之一,涉及所有儿童期胚胎性肿瘤,如肺癌、卵巢癌、乳腺癌、肝癌及其他消化道肿瘤。近期研究表明,正常结肠上皮有 IGF2 印记缺失的人都患有 LOI 相关的结肠癌[15]。但由于在病变周围的非瘤组织中或癌前病变的组织中也可检出 LOI,因此认为,LOI是肿瘤发生中的前期事件,它可以增高人和大鼠肠癌发生的危险性[16]。
参考文献
公司简介


微信号|一米生物
服务热线|400-097-3606
