
随着低起始量染色质免疫共沉淀技术测序(Chromatin Immunoprecipitation sequencing,ChIP-seq)方法的开发,早期胚胎细胞数量的限制被逐渐突破,有关哺乳动物早期胚胎发育过程中组蛋白修饰重编程的研究不再局限于免疫荧光染色等观察手段[1-3],关于早期胚胎发育各阶段的各类组蛋白修饰的报道也越来越多。
2016 年,多个研究团队同时在 Nature 等杂志上发表论文,首次报道了小鼠早期胚胎中多种组蛋白修饰的变化图谱,这些研究显示,在受精后,父源基因组中的 H3K4me3 会被迅速去除,但是在 majorZGA 阶段这一标记则被重新建立。相比之下,人们在母源基因组中发现了 H3K4me3 的非传统形式(ncH3K4me3),其在启动子和远端区域中都有宽的信号[1241ncH3K4me3 很容易在成熟卵母细胞中建立,直到 majorZGA 阶段才被经典的 H3K4me3 取代。赖氨酸去甲基化酶 Kdm5a 和 Kdm5b 具有主动去除宽的 H3K4me3 信号的作用,这对于 ZGA 和早期胚胎发育而言必不可少,但是在卵母细胞阶段,Kdm5b 过表达导致转录组重新激活,表明 ncH3K4me3 可能是维持卵母细胞全基因组沉默状态的重要原因[2]。
另一方面,基因启动子区域的传统的 H3K4me3 信号的宽度在小鼠植入前胚胎发育过程中是高度动态的,并且与基因表达水平呈正相关。H3K4me3 信号宽度的动态性可能为早期胚胎中的表观遗传调控提供了一种新的机制。研究还发现较宽的 H3K4me3 信号(>5kb)在早期胚胎中含量远高于其他类型细,并且这些宽的 H3K4me3 信号多富集在谱系特异性的基因上。敲降 Kdm56 可以干扰启动子 H3K4me3 信号的宽度并对胚胎发育产生严重的影响,这表明宽的启动子 H3K4me3 标记可能有助于谱系特异性因子的稳定表达,与细胞命运的维持息息相关[1]。
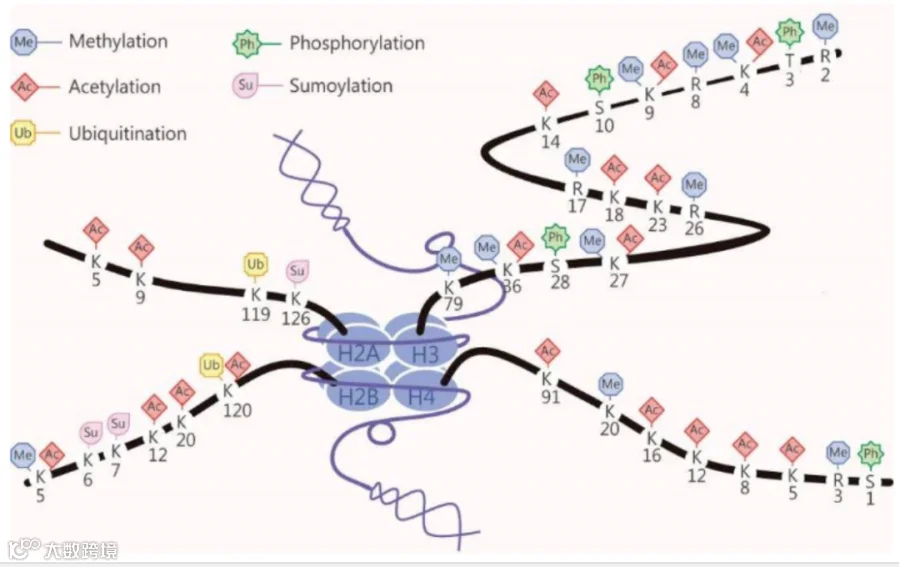
图1 组蛋白修饰示意图
H3K27me3 常定位于启动子区域,起到抑制基因表达的作用。在小鼠早期胚胎发育过程中,早在 PN5 时期母源和父源基因组启动子区域中的 H3K27me3 修饰都已出现广泛的丢失,随后在从桑葚胚期到囊胚期的过渡过程中出现快速动态变化[1,4,5]。另一方面,母源 H3K27me3 介导的基因印迹也在早期小鼠胚胎中发现。H3K27me3 印迹可能是在卵子发生期间建立,并在植入前的胚胎中维持的。其在内细胞团中被稀释,且大多数 H3K27me3 介导的印迹在 E6.5 胚胎的外胚层干细胞中消失。最近的研究表明,卵母细胞来源的 H3K27me3 介导的印迹在植入后将转变为 DNA 甲基化依赖性的印迹,其中母源因子 Eed 和合子表达的 Dnmt3a/b 都可能参与其中,这也反映了 H3K27me3 和 DNA 甲基化在控制印迹中的互补作用[4-6]。
二价修饰为分化后的转录激活提供了发育调控因子[7]。小鼠植入前胚胎中的二价基因数量远低于胚胎干细胞,这表明在稳定的细胞系中二价修饰的存在比在过渡阶段更重要。值得一提的是,PRC2 的核心成分 Ezh2 和 Suz12 负责 H3K27me3 的建立,它们在胚胎中靶向大多数可遗传的二价基因。这一发现可能有助于我们理解在早期胚胎发育中从全能性退出到分化后 PRC2 介导的 H3K27me3 的调控模式[1]。清华大学的颉伟团队在对植入后胚胎的研究中发现,在 E6.5 的外胚层干细胞中,发育相关基因的启动子区域存在一类更强的二价基因,即“超二价基因”,这在 E7.5 胚胎的相应结构中也很明显,但在 E6.5 内脏内胚层,小鼠胚胎干细胞和体细胞中却并没有这种现象。Kmt2b 在 E6.5EPI 的超二价性中起着至关重要的作用,而某种补偿机制则参与了后来的胚胎发育,此外这个工作还证明 DNA 低甲基化参与了二价的维持[8]。以上结果揭示了植入前胚胎中独特的组蛋白修饰模式和调控机制[9]。
参考文献
扫码关注获得更多内容

小红书@一米生物

视频号@一米生物

知乎@一米生物
公司简介


微信号|一米生物
服务热线|400-097-3606
