
体细胞核移植(SCNT)是将体细胞移植到去核的卵母细胞中,并启动发育的过程,是目前体外获得完整全能性的重要手段。但是,体细胞核移植中的重编程效率低下、胚外组织的缺陷以及克隆个体的异常限制了其应用。近年来,关于体细胞核移植中多种表观遗传障碍的报道越来越多,为科研人员探索提高重新编程效率提供了重要线索。
1. DNA 甲基化的表观遗传障碍
体细胞通常具有较高的 DNA 甲基化水平[1]。研究表明,胚胎激活后卵母细胞的 TET3 会迅速进入重组胚胎的假性原核(pseudopronucleus,PPN)催化从 5-甲基胞嘧啶到 5-羟甲基胞嘧啶的转换,说明在 SCNT 胚胎发育过程中 TET3 发生了主动去甲基化,这与正常胚胎发育相似[2]。全基因组亚硫酸氢盐测序(WGBS)表明,到囊胚阶段,核移植胚胎已表现出与正常受精囊胚相似的非常低的 DNA 甲基化水平,表明体细胞核移植胚胎成功实现了全基因组水平的 DNA 去甲基化[3]。但是,在小鼠 SCNT 胚胎发育到 1-细胞后期时,这种去甲基化事件还没有完成[4]。在 2-细胞和 4-细胞时期,体细胞核移植胚胎的 DNA 甲基化水平普遍高于相应时期的自然受精胚胎。在小鼠 4-细胞期体细胞核移植胚胎中,尤其是在出现了发育停滞的样本中,基因体区域的平均甲基化水平显著增加,与供体细胞的趋势更为相似。而在囊胚阶段,虽然实现了整体的DNA 去甲基化,但是在局部的调控元件上,甚至在植入后仍然可以检测到异常的 DNA 甲基化模式[5],这些 DNA 甲基化的异常可能是导致核移植动物出生率低的原因之一。
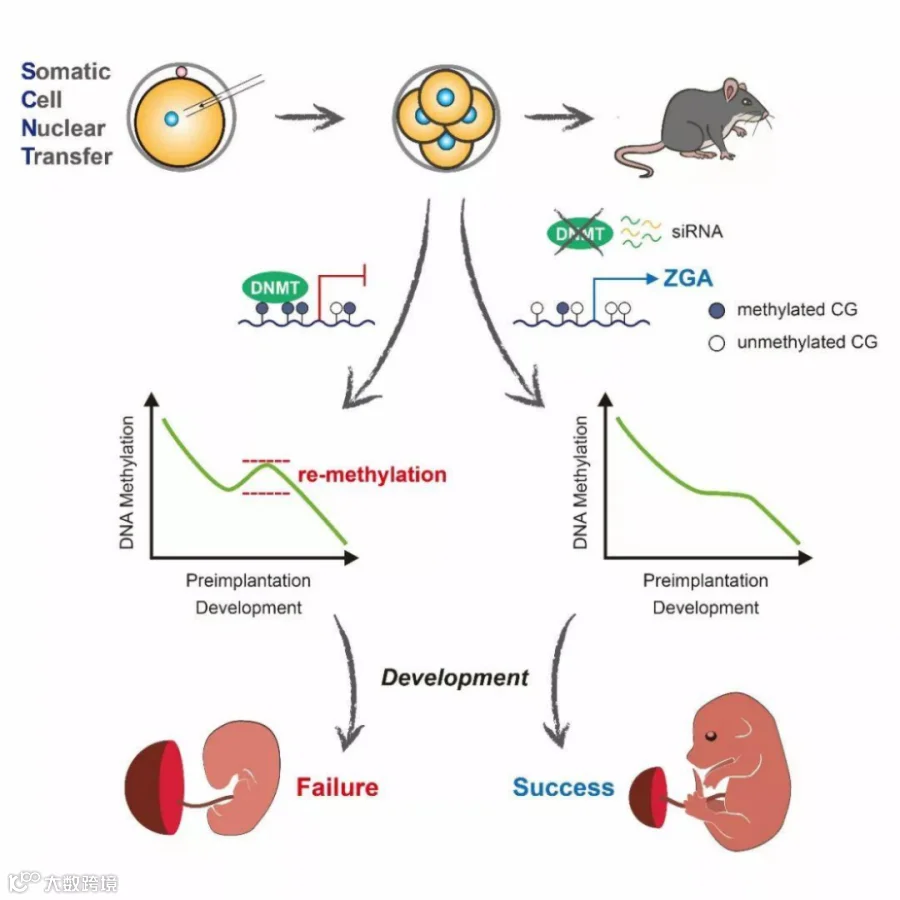
图1 抑制异常的 DNA 再甲基化可改善体细胞核移植胚胎的植入后发育
2. 组蛋白修饰的异常
2014年,哈佛大学的张毅团队发现与体外受精 2-细胞胚胎相比SCNT 2-细胞胚胎中有 222 个重编程抵抗区未能被激活,且这些重编程抵抗区在体细胞中富含 H3K9me3 修饰。通过在卵母细胞中异位表达 H3K9me3 特异性去甲基酶 Kdm4d 或在供体小鼠胚胎成纤维细胞中敲除 H3K9 甲基转移酶 Suv39h1 和 Suv39h2 去除这种表观遗传标记,可以显著提高核移植胚胎的重编程效率[6]。后来,高绍荣团队通过单细胞测序的手段发现 H3K9me3 修饰的异常是核移植胚胎 2-细胞阻滞的重要原因,通过显微注射 Kdm4bmRNA有助于 SCNT 胚胎突破 2-细胞阻滞,并最终显著提高囊胚率、核移植胚胎干细胞建系效率,甚至出生率[5]。另有研究证明供体细胞中的 H3K9me3 还可妨碍核移植胚胎拓扑相关结构域的去除[7]。这些发现表明,H3K9me3 修饰无法去除是核移植胚胎重编程的重要障碍。
组蛋白 H3K4me3 修饰的正确去除与重建在核移植胚胎发育过程中也至关重要。单细胞测序的结果显示,Kdm5b 在 4-细胞阻滞核移植胚胎中未能被激活,而 Kdm5b 的过表达有助于核移植胚胎跨越 4-细胞阻滞,并显著提高囊胚的形成率和质量[5]。考虑到 Kdm5b 作为H3K4me3 去甲基化酶的作用以及 H3K4me3 在转录起始中的作用,核移植胚胎中供体特异性 H3K4me3 标记可能是其重编程的障碍,目前这一观点已经在非洲爪蟾、人类和牛的核移植胚胎中得到证实[8,9]。
核移植胚胎中,H3K27me3 修饰同样存在明显的缺陷,而这种缺陷主要表现在对印迹基因的调控上[3,10,11]。目前有 70 余个基因被发现具有父源特异性 DNA 酶I超敏位点且不具有 DNA 甲基化,但却同时含有母源特异性 H3K27me3 修饰。而 Gab1、Sfmbt2 和Slc38a4 等与甲基化无关的印记基因均在其中[12],因此,人们推测 H3K27me3 介导的印迹缺陷可能导致核移植胚胎胎盘异常。而相应的事实证据也在核移植小鼠胚胎植入后阶段被发现,多项研究证明在核移植胚胎中丢失 H3K27me3 印迹会干扰小鼠植入后的发育,而到目前为止这种缺陷可以最早在囊胚期胚胎中被发现[13]。除 H3K27me3 印迹缺失对核移植胚胎植人后的影响外,另一组研究表明体细胞 H3K27me3 修饰的残留同样也是核移植胚胎发育的障碍。H3K27me3 特异性去甲基化酶 Kdm6a 的过表达显著增加了核移植胚胎的囊胚形成率[14]。总体而言,尽管其机制尚不清楚,富集在特定区域(如印迹基因)的 H3K27me3 修饰及其适当去除对核移植胚胎重编程的成功是至关重要的。
组蛋白乙酰化的重建对于克隆胚胎中合子基因的激活至关重要。组蛋白去乙酰化酶抑制剂能显著提高组蛋白乙酰化水平和克隆成功率,这在核移植中已得到广泛应用[15]。另一方面,一些乙酰化标记,如 H4K8ac 和 H4K12ac,在核移植胚胎发育过程中持续存在于基因组中,这可能是导致克隆效率低下的原因。最近一项工作绘制了在核移植胚胎发育过程中 H3K9ac 全基因组图谱,并从中发现异常的乙酰化区域妨碍了合子基因组的激活[16]。
3. 染色质可及性与染色质高级结构的影响
小鼠核移植胚胎发育过程中染色质可及性的完整模式尚待阐明,但最近的一项研究分析了供体细胞和 1-细胞晚期小鼠核移植胚胎中的 DNaseI 高敏位点(DNase I hypersensitive site,DHS)。他们发现,核移植胚胎的染色质可及性重编程在激活后 12 小时内基本已完成,供体细胞的 DNaseI 高敏位点在 12 小时内发生了剧烈变化并在一定程度上重现了体外受精受精卵的 DNaseI 高敏位点。这种变化是不依赖 DNA 复制的,在诸如爪蟾等模式动物的核移植胚胎中是保守的[17]。同济大学高绍荣团队最近的一项研究描述了核移植早期胚胎 3D 染色质结构的时空动态,揭示了 3D 染色质结构在核移植后可以迅速重组为自然受精胚胎样状态的发育事件。然而,在植入前核移植胚胎发育过程中,异常的拓扑相关结构域和 A/B 区室结构仍可以被观察到[7,18]。
参考文献
扫码关注获得更多内容

小红书@一米生物

视频号@一米生物

知乎@一米生物
公司简介


微信号|一米生物
服务热线|400-097-3606
