
1. 染色质重塑因子在重编程中的作用
目前的研究显示,多种染色质重塑因子通过其染色质重塑活性来调节体细胞重编程。在染色质重塑复合物家族中,已证明 esBAF(胚胎干细胞中的 Bmm /Brg 相关因子)和 Ino80 参与了诱导多能干细胞重编程。这些重塑复合物介导开放染色质结构的产生,并被认为在重编程期间促进 OSKM 因子的结合与转录活性[1,2]。另外,CHD 家族重塑因子 Chd1 的下调会导致异染色质的积累,并伴随着重编程效率的显著隆低[3,4]。相反,NuRD 复合物包含组蛋白去乙酰化酶活性。在缺乏该复合物的核心亚基 Mbd3 时,胚胎干细胞表现出与白血病抑制因子无关的自我更新能力提高。有趣的是,即使在没有 c-Myc 或 Sox2 的情况下,Mbd3 的消耗也会显著提高诱导多能干细胞的获得效率[5]。一个可能的解释是 Mbd3/NuRD 通过与 OSKM 转录因子直接相互作用募集到下游 OSKM 靶基因,并在诱导多能干细胞诱导过程中抵消它们的重新激活,虽然在胚胎干细胞中的相关研究已日臻完善[6,7],但是在诱导多能干细胞重编程过程中的更确凿的证据和深入的分子机制研究则有待填补。
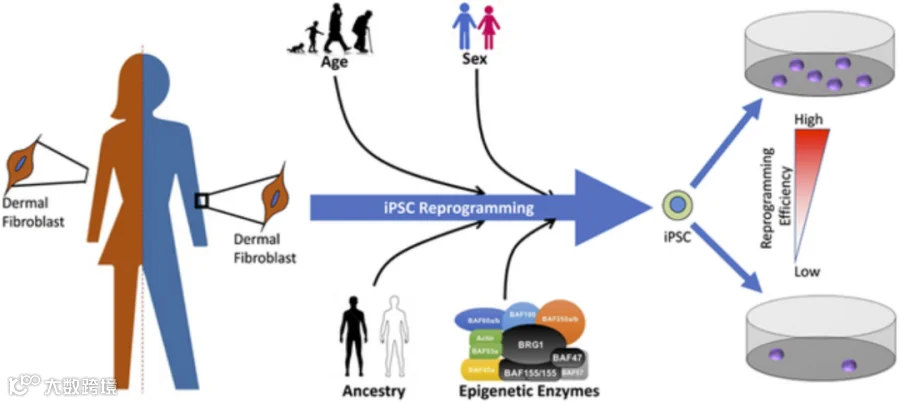
图1 调节人类 iPSC 重编程效率的因素[2]
2. 组蛋白修饰与组蛋白变体在重编程中的作用
诱导多能干细胞中各类组蛋白修饰对应的功能与胚胎干细胞中的具有一定程度的相似性,但诱导多能干细胞重编程过程中存在着的一系列特殊的组蛋白修饰及组蛋白变体,并且对重编程过程的发生起到关键的调控作用。
H3K4me2 分布在全基因组上的快速变化是在重编程过程中观察到的最早事件之一,通过染色质免疫共沉淀测序技术人们观测到 H3K4me2 信号峰在超过一千个基因的启动子和增强子区域表现出巨大变化,而这些基因中包括许多多能性相关基因和发育调控基因座。H3K4me2 往往被认为是转录激活的标志之一,Wdr5 是负责 H3K4 甲基化的 Set/MLL 组蛋白甲基转移酶复合物的关键成分,它可以直接与 OCT4 相互作用[8]。这种相互作用可能解释了在诱导多能干细胞重编程早期 OCT4 结合位点 H3K4 甲基化的快速获得,也可能是体细胞命运逆转的关键诱因之一。与含激活性组蛋白修饰标志物 H3K4me2/3 的区域形成鲜明对比的是,富含抑制性 H3K9me3 修饰的染色质结构域是不利于初始 OSKM 的结合的,而通过下调甲基转移酶 Suv39H1/2 降低 H3K9me3 水平即可增强这些区域的 OCT4 和 SOX2 结合并提高重编程效率[9],因此,H3K9me3 标记的异染色质区域被认为是体细胞重编程过程中的表观遗传屏障。另外,其他的重要组蛋白修饰,如 H3K27me3 和 PRC2 之间的调控网络也影响着体细胞重编程的效率。
组蛋白变体的变化在重编程过程中也发挥重要作用。在早期的研究中人们发现,组蛋白分子伴侣 Asf1a 的过表达有利于维持胚胎干细胞的多能性,并增强了成纤维细胞(hADFs)的诱导多能干细胞诱导效率。ASF1 与组蛋白 H3-H4 异二聚体非选择性结合,并促使其从细胞质中导人细胞核,从而直接调节 H3-H4 二聚体中组蛋白 H3.1/2 和 H3.3 的转换[10]。而 Th2a 和 Th2b 的联合过表达也可使诱导多能干细胞诱导效率明显提升,在这种方式获得的诱导多能干细胞细胞系中可观察到 DNase Ⅰ 超敏位点的增多,表明染色质开放性的增加[10,11],这与体细胞重编程初期染色质趋于开放的现象是一致的。
3. 体细胞重编程过程中的异常甲基化
在过去的研究中,人们利用相同遗传背景的细胞制备了诱导多能干细胞系与胚胎干细胞系,并比较了其 DNA 甲基化组和转录组差异。与胚胎干细胞相比,诱导多能干细胞系存在大量异常 DNA 甲基化区域;这说明了诱导多能干细胞重编程过程中基因表达的异常与 DNA 甲基化之间存在明显的相关性[12],诱导多能干细胞重编程过程中的不完全去甲基化可能是影响其细胞命运逆转的关键原因。在后续的研究中,人们证明了维持型 DNA 甲基转移酶 DNMT1 对诱导多能干细胞诱导效率有较大影响,通过敲除或敲降 DNMT1 可造成全基因组水平的 DNA 甲基化减少,并大幅提高了诱导效率,而从头甲基转移酶 Dnmt3a 和 Dnmt3b 则对体细胞重编程及命运逆转并未有明显影响[13]。而 TET 家族,尤其是 TET1 则被证明参与 OCT4 和 NANOG 的全能性调控网络,并与 OCT4 和 TET1 的启动子周围 DNA 去甲基化的正反馈相关[14,15]。
参考文献
扫码关注获得更多内容

小红书@一米生物

视频号@一米生物

知乎@一米生物
公司简介


微信号|一米生物
服务热线|400-097-3606
