
端粒调控
衰老是一个不断发展变化的退行性过程,常伴有组织干细胞耗竭、组织炎症、基质改变、细胞衰老和代谢功能障碍等。这些细胞和组织的变化反映了线粒体、蛋白稳态、细胞间通信、营养感应、表观遗传学和 DNA 修复等潜在的异常变化,这些异常变化导致了基因组不稳定性和损坏,包括端粒功能障碍。随着对各种衰老相关分子机制更深入的了解,人们意识到端粒功能障碍是驱动衰老及其相关疾病的分子通路的诱因或促进剂。端粒由重复的核苷酸序列组成,形成“帽子结构”,发挥维持染色体完整性的作用。人的端粒维持相关基因发生缺陷时,会引起生殖细胞和体细胞的退行性疾病,如先天性角化病、特发性肺纤维化、溃疡性结肠炎等。端粒等位基因功能丢失或可诱导的基因敲除小鼠加快了端粒功能障碍和衰老、早衰综合征、慢性炎症以及退行性疾病等相关性的研究[1]。内源性端粒酶的重新激活可逆转具有端粒功能障碍小鼠的过早衰老。鳉鱼和斑马鱼都是端粒生物学研究的模型,它们的端粒长度与人类相似,而其端粒功能障碍的表型同啮齿类动物模型更相似些。端粒和端粒酶在衰老早衰综合征、年龄相关退行性疾病(如神经退变和癌症)中的作用似乎无处不在。
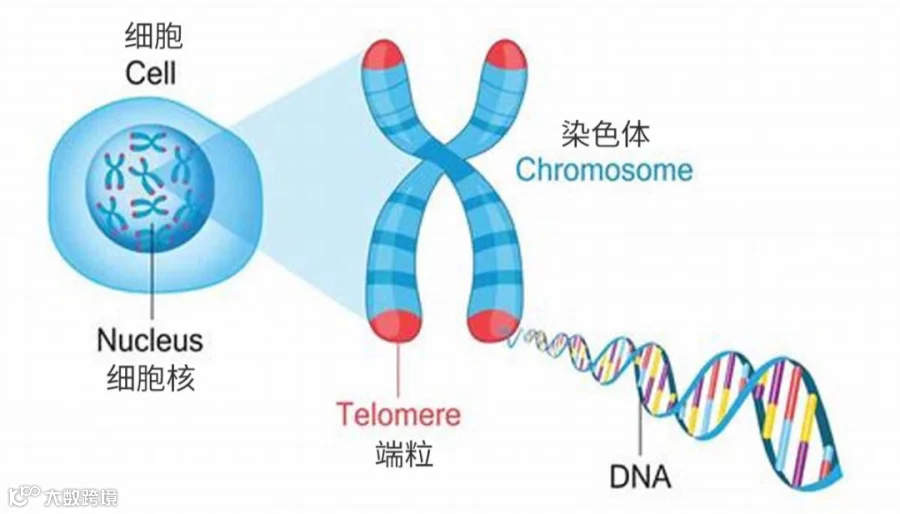
图1 端粒调控衰老的发生
DNA 甲基化
DNA 甲基化作为主要的表观遗传修饰方式之一,在衰老及衰老相关疾病中起到重要作用,在某些特定位置 DNA 甲基化修饰随着生物年龄增长产生或消失。因而,DNA 的甲基化修饰变化提供了一个衡量生理年龄的标志。DNA 甲基化主要是选择性地将甲基添加到基因组 DNA 序列中特定的区域,通常是 CpG 岛发生甲基化,而 CpG 岛区域经常定位在基因的启动子区域,对调控基因的表达、转座子沉默、可变剪切、基因组稳定性维持至关重要[2]。最为经典的是胞嘧啶上的甲基化修饰,胞嘧啶可以被甲基化形成 5-甲基胞嘧啶,通过对双生子及新生儿与百岁老人 CD4+T 淋巴细胞研究,表明衰老进程中 DNA 甲基化修饰比较分散,这主要是由于环境因素导致的表观遗传漂移或者基因组自身所发生的错误。DNA 甲基化对于衰老进程的影响可以通过对特定基因的表达调控实现,POLG 突变小鼠会出现加速衰老的表型,而在正常情况下 POLG 的表达会受到其启动子区甲基化水平的调控,这种甲基化的状态受到体内炎症因子的影响。
衰老进程中全基因组呈现低甲基化状态并不代表所有位点的低甲基化。在人及动物模型动脉粥样硬化中,发现全基因组 DNA 低甲基化,但抗动脉粥样硬化基因 ESR1/2、ABCA1、KLF4 的驱动子区域呈现超甲基化状态[3]。多梳家族蛋白目的基因在衰老及癌症中呈现出超甲基化状态。多梳家族蛋白形成的复合物与 DNA 稳定、染色质重塑及转录抑制作用相关。Dnmt3a/b-/- 小鼠胚胎干细胞中,多梳家族蛋白会倾向于与未甲基化的 CpG 岛区域结合,调控甲基化水平[4]。多梳家族蛋白识别未甲基化的 DNA 的机制是通过 KDM2B 蛋白去招募多梳抑制蛋白复合物 1/2 实现的。此外,DNMT1 活性在衰老过程中会出现持续性的下降。另外,DNMT3a/3b 缺失的造血干细胞移植实验中发现,这种缺陷的造血干细胞向髓系细胞和淋系细胞分化能力不变,但是增殖能力减弱,证明了 DNMT3a/3b 会保护造血干细胞的自我更新能力[5]。在 DNMT3a 敲除的神经干细胞中,神经元发生相关基因(如 Dlx2、Sp8 和 Neurog2)出现下调,与 H3K27me3 修饰水平升高正相关,揭示了 DNA 甲基化与组蛋白甲基化修饰协同调节靶基因的表达[6,7]。
参考文献
扫码关注获得更多内容

小红书@一米生物

视频号@一米生物

知乎@一米生物
公司简介


微信号|一米生物
服务热线|400-097-3606
