
上皮间质转化(epithelial-mesenchymal transition, EMT)是指上皮细胞通过特定程序转化为具有间质表型细胞的生物学过程。通过 EMT,上皮细胞失去了细胞极性,失去与基底膜的连接等上皮表型,获得了较高的迁移、侵袭、抗凋亡和降解细胞外基质的能力等间质表型。EMT 是上皮细胞来源的恶性肿瘤细胞获得迁移和侵袭能力的重要生物学过程。
多种蛋白参与 EMT,其中 E-钙黏蛋白(E-cadherin, E-cad)对于正常的上皮细胞间连接稳定性非常重要,其表达水平与 EMT 的发生以及肿瘤的侵袭能力呈负相关;Vimentin 和 N-cadherin 的表达水平与 EMT 的发生以及肿瘤的侵袭能力呈正相关;转录因子 SNAI1/Snail、SNAI2/Slug、ZEB1、ZEB2 和 Twist 等可下调 E-cad 的表达而促进 EMT 的发生,继而促进肿瘤的转移[1]。
EMT 是上皮细胞失去极性转换成为具有侵袭和迁移能力的间质细胞的过程,参与细胞重编码[2],在胚胎发育过程中与干细胞的多能性[3]也密切相关。因此,EMT 对肿瘤干细胞的干性有增强作用,或者说肿瘤干细胞具有 EMT 特性[1]。
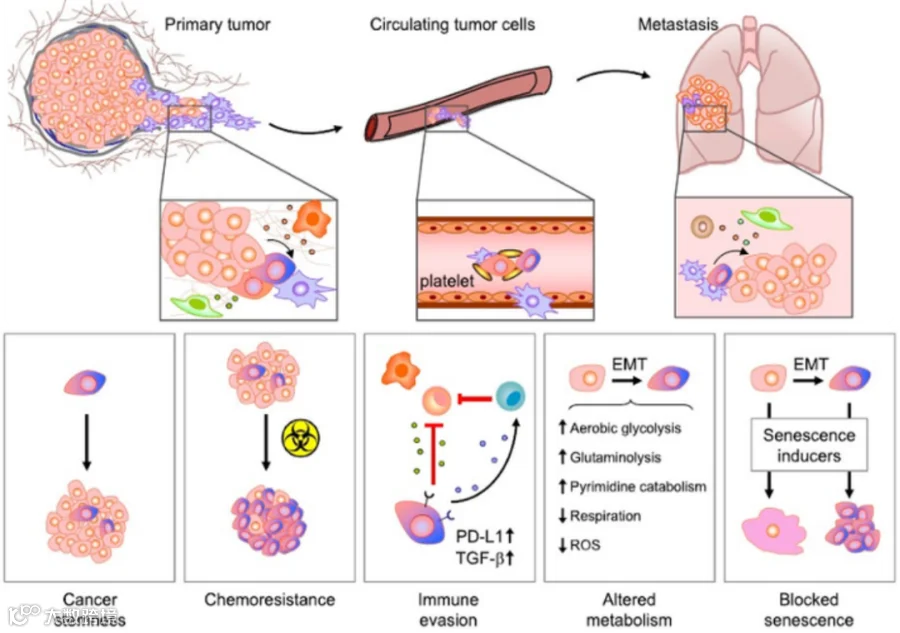
图1 肿瘤转移途径:上皮间质转化
在过去的 20 多年时间里,肿瘤细胞 EMT 的活化一直被认为是导致肿瘤转移形成的诸多复杂过程中的第一步,是肿瘤细胞从原位肿瘤侵袭至周围基质组织的主要作用机制。但随着研究不断地深入,EMT 在肿瘤转移中参与调控的生物进程绝不仅仅局限在侵袭这一步,EMT 的活化还能赋予上皮性肿瘤细胞多种肿瘤干细胞特性[4,5]。例如在乳腺癌中,这些特性包括肿瘤干细胞的特异细胞表面标记物 CD44 表达增加和 CD24 的表达下调,悬浮培养条件下成球能力的显著升高以及荷瘤小鼠模型中肿瘤形成能力的显著增强。
除此之外,多种人肿瘤类型的体内连续成瘤实验也证实了肿瘤细胞 EMT 的活化并促使细胞转化进入肿瘤干细胞状态。这些研究成果有力地说明 EMT 进程在驱动肿瘤细胞恶性转变时所起作用的多样性,同时也体现了 EMT 在整个肿瘤发生发展中的重要作用[6]。
EMT 进程与肿瘤干细胞之间的联系表明,EMT的活化能够使得非肿瘤干细胞转化成为肿瘤干细胞。另外,肿瘤干细胞具有与成体干细胞相似的多能分化特性,可以分化成为非肿瘤干细胞,而 EMT 的逆向过程 MET 的发生很有可能引起这种分化的出现。因此肿瘤细胞中 EMT 和 MET 的相互转化也从另一方面论证肿瘤细胞具有极强的可塑性[7,8]。
肿瘤干细胞具有生成肿瘤的能力,散播至远端位点的肿瘤细胞就像是“种子”一样有能力形成肿瘤转移灶。而诸多研究证实 EMT 进程驱使肿瘤细胞进入干细胞样阶段。因此在肿瘤转移中,EMT 的活化不仅能将肿瘤细胞从原位组织中解离,还能赋予远端位点散播的肿瘤细胞干细胞特性,促使肿瘤远端转移灶的形成。肿瘤细胞中 EMT 的活化能够导致大量肉眼可见的肺部转移出现,如果肿瘤细胞被人为注射入静脉循环系统,它们就可以直接跳过侵袭和外渗的转移步骤[9]。
Robert A. Weinberg 等科学家在研究肺组织中浸润的肿瘤细胞与细胞外基质蛋白之间关系时发现,那些容易在转移灶“扎根”的肿瘤干细胞样细胞比其他细胞拥有富集更多由整合素聚集形成的成熟黏附斑块[10,11]。成熟黏附斑块能够强有力地激活与肿瘤干细胞样细胞增殖密切相关的黏着斑激酶 FAK,从而提升肿瘤细胞的“扎根”能力[12]。
后续研究发现造成细胞亚群之间不同黏附斑块的出现主要是由于肿瘤细胞外渗入组织后细胞突出物延伸能力不同。后者是由肌动蛋白和整合素聚合而成,被称为“伪足样突起(filopodium-like protrusions, FLPs)”。FLPs 只有在 EMT 活化的细胞中才会大量形成,进而建立整合素-胞外基质联系以便黏附斑块发展,进而强有力地激活 FAK 信号通路。值得注意的是,在多种肿瘤模型中,人为激活 EMT 进程均会导致细胞 FLPs 延伸能力的显著增强。而 FLPs 延伸能力的增加会促使肿瘤细胞能更有效率地在转移位点生存、增殖以及最终形成肿瘤转移灶[13]。EMT、FLPs 以及 FAK 之间的关系很好地为 EMT 促进肿瘤转移灶的形成提供了另外一种作用机制[14]。
参考文献
公司简介


微信号|一米生物
服务热线|400-097-3606
