
成纤维细胞
肿瘤组织中位于肿瘤上皮细胞附近的成纤维细胞对于肿瘤发生发展非常重要,同时也对肿瘤干细胞的干性维持有促进作用。
首先,肿瘤干细胞本身可以分泌一些特异的生长因子影响其附近的成纤维细胞从静息或者不活跃状态转变为活跃状态,并使得这些处于活跃状态的肿瘤相关成纤维细胞(CAF)分泌维持肿瘤干细胞干性所需要的蛋白。例如,有研究表明,CAF 可以通过旁分泌方式增加肝脏肿瘤干细胞的数量,其中分泌的肝细胞生长因子( hepatocyte growth factor, HGF ),能激活 c-Met/FRA1/HEY1 信号,从而使肝癌细胞发展 CAF 依赖性的HCC[1]。在头颈部鳞状细胞癌中,最近发现 CAF 分泌的骨膜蛋白能显著增强头颈部鳞状细胞癌的肿瘤干细胞样表型、增殖和侵袭。通过这种机制,骨膜蛋白与肿瘤干细胞膜上的蛋白酪氨酸激酶7 (PTK7)结合,并激活下游 Wnt/β-Catenin 信号通路,以促进肿瘤干细胞的表型[2]。在前列腺癌中,雄激素受体也可被前列腺 CAF 分泌的干扰素-γ(IFN-γ)和巨噬细胞集群体刺激因子(M-CSF)激活,从而促进前列腺癌细胞中干细胞标志物的表达,获得前列腺癌干细胞样细胞的表征及功能[3]。
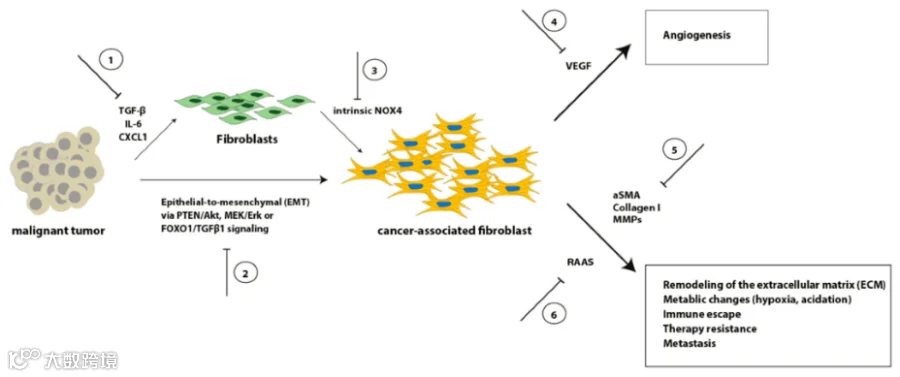
图1 成纤维细胞向CAF转化
血管内皮细胞
就像肿瘤相关成纤维细胞一样,肿瘤组织中的血管内皮细胞也能促进肿瘤干细胞的功能及表型。有报道称,肝脏组织中的血管内皮细胞可通过激活 Nanog 逆转录基因P8 (NANOGP8)通路,以旁分泌方式介导结直肠癌中肿瘤干细胞的激活[4]。此外,来自肿瘤微血管内皮细胞的条件培养基可以恢复脑胶质瘤的肿瘤干细胞表型,这是由条件培养基中存在的一种主要可溶性因子 bFGF 介导的[5]。
免疫细胞
很多研究主要针对肿瘤细胞本身找原因,虽然肿瘤细胞基因组的突变和不稳定性等是肿瘤的重要特性,但研究发现肿瘤微环境与肿瘤细胞相互作用,对肿瘤细胞的各种特性起到至关重要的调节作用。其中微环境中的免疫细胞似乎起着更重要、不可替代的作用。
肿瘤干细胞与肿瘤组织中的巨噬细胞、髓源性抑制细胞和 T 细胞相互作用[6],一方面,肿瘤干细胞能分泌细胞因子和趋化因子,招募并极化肿瘤相关巨噬细胞(TAMs)。这些 TAMs 可以来自骨髓的巨噬细胞(BMDMs),也可是局部组织中的巨噬细胞(包括脑中的小胶质细胞、肝中的 Kupffer 细胞和肺中的肺泡巨噬细胞)。
另一方面,TAMs 可以支持肿瘤干细胞的干性获得及维持。同样,肿瘤干细胞可以通过分泌可溶性因子和外泌体产生趋化因子(如 CCL2 和 CCL5 招募骨髓中产生的髓源性抑制细胞到肿瘤组织中,促进髓源性抑制细胞的浸润、扩张和激活,而髓源性抑制细胞被招募到肿瘤组织中后,促进肿瘤干细胞的干性维持。肿瘤干细胞与 T 细胞的相互作用机制也与上述两种免疫细胞相似。
图2 肿瘤干细胞与免疫细胞的相互作用
以前列腺癌为例,一方面,临床上经典抗雄治疗促进前列腺肿瘤干细胞的富集,而后者能引起免疫细胞功能的变化及免疫因子的产生水平。在体外,抗雄处理小鼠前列腺肿瘤细胞 TRAMP-C2,能促使后者细胞表面 MHC-I 和 Fas 的表达升高,从而提高 TRAMP-C2 细胞对 T 细胞调节的细胞杀伤作用的敏感性[7]。同时,抗雄治疗能通过调节肿瘤细胞上 AR 和抗凋亡基因 NAIP(NLR 家族,神经元凋亡抑制蛋白)的表达增强人前列腺癌细胞系对 T 细胞调节的细胞杀伤作用的敏感性[8]。
另外,抗雄治疗能激活胸腺的再生,从而增加外周 T 细胞的绝对数量,尤其是 CD4+T 细胞,并提高细胞毒性 T 淋巴细胞(cytotoxic T lymphocytes,CTL)的活性[9,10]。最后,一项临床前研究说明,经过前列腺特异抗原设计的肿瘤疫苗处理后,抗雄治疗能消除 CD4+T 细胞对前列腺肿瘤特异抗原的耐受性,从而促进 CD4+T 细胞的扩增和效应细胞因子的分泌[11]。抗原特异肽段和 MHC-Ⅱ 分子的复合物与 naive CD4+T细胞表面 TCR 结合后能诱导后者向效应 T 细胞和长期留存的记忆 T 细胞极化,增强免疫反应。
另一方面,抗雄治疗后扩增的免疫细胞也能促进前列腺癌去势抵抗(CRPC)及前列腺肿瘤干细胞的形成。例如,据临床病例分析,接受抗雄治疗后的前列腺癌病人中 CD3+ 和 CD8+T 淋巴细胞,CD68+ 巨噬细胞和 CD56+ 自然杀伤性细胞(natural killer cells,NK cells)的比例均显著升高[12]。相似地,对前列腺癌小鼠模型的分析显示,抗雄治疗后,肿瘤微环境中侵润的多种免疫细胞的数量和比例也都大幅升高(包括 T 细胞,B 细胞,NK 细胞和髓系细胞),且 B 细胞可通过分泌淋巴毒素激活肿瘤细胞中的 IKK-α 和 STAT3,进而促进 CRPC 及前列腺肿瘤干细胞的形成[13]。
另外,大量研究表明,炎症因子可促进 CRPC 及前列腺肿瘤干细胞的发生和进展。IL-6 可通过激活 SRC-1,调控 GRB2,SHC 和 JAK-1 的表达以及抑制最终会逐步发生 CRPC[14]。IL-4 可作用于 AR 共激活因子,如 CBP/P300 和 NF-κB,促进 CRPC 的进展[15]。IL-8 在 CRPC 中过表达,能与 NF-κB 相互作用且可通过激活 SRC 和 FAK 等信号通路促进 CRPC 及前列腺肿瘤干细胞的产生和肿瘤转移[16]。CXCR 家族分子 CXCR4、CXCR2/CXCR3、CXCR6 和 CXCR7,也可通过相关炎症信号通路促进 CRPC 及肿瘤干细胞的发生和进展[17,18,19]。
参考文献
公司简介


微信号|一米生物
服务热线|400-097-3606
