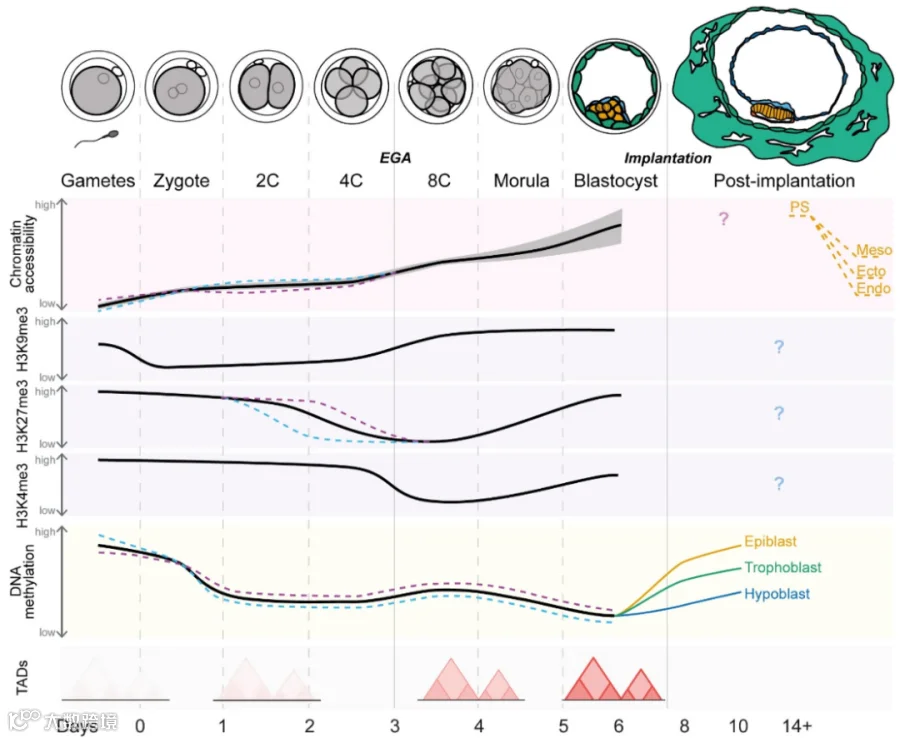
[1] Xu Q, Xie W. Epigenome in Early Mammalian Development: Inheritance, Reprogramming and Establishment. Trends Cell Biol. 2018;28(3):237-253. doi:10.1016/j.tcb.2017.10.008
[2] Yeste M, Jones C, Amdani SN, Coward K. Oocyte Activation and Fertilisation: Crucial Contributors from the Sperm and Oocyte. Results Probl Cell Differ. 2017;59:213-239. doi:10.1007/978-3-319-44820-6_8
[3] Minami N, Suzuki T, Tsukamoto S. Zygotic gene activation and maternal factors in mammals. J Reprod Dev. 2007;53(4):707-715. doi:10.1262/jrd.19029
[4] Jukam D, Shariati SAM, Skotheim JM. Zygotic Genome Activation in Vertebrates. Dev Cell. 2017;42(4):316-332. doi:10.1016/j.devcel.2017.07.026
[5] Schulz KN, Harrison MM. Mechanisms regulating zygotic genome activation. Nat Rev Genet. 2019;20(4):221-234. doi:10.1038/s41576-018-0087-x
[6] Zhang Y, Xiang Y, Yin Q, et al. Dynamic epigenomic landscapes during early lineage specification in mouse embryos. Nat Genet. 2018;50(1):96-105. doi:10.1038/s41588-017-0003-x
[7] Yao C, Zhang W, Shuai L. The first cell fate decision in pre-implantation mouse embryos. Cell Regen. 2019;8(2):51-57. Published 2019 Nov 20. doi:10.1016/j.cr.2019.10.001
[8] Legault LM, Bertrand-Lehouillier V, McGraw S. Pre-implantation alcohol exposure and developmental programming of FASD: an epigenetic perspective. Biochem Cell Biol. 2018;96(2):117-130. doi:10.1139/bcb-2017-0141
[9] Yu B, Smith TH, Battle SL, Ferrell S, Hawkins RD. Superovulation alters global DNA methylation in early mouse embryo development. Epigenetics. 2019;14(8):780-790. doi:10.1080/15592294.2019.1615353
[10] Smith ZD, Chan MM, Mikkelsen TS, et al. A unique regulatory phase of DNA methylation in the early mammalian embryo. Nature. 2012;484(7394):339-344. Published 2012 Mar 28. doi:10.1038/nature10960
[11] Inoue A, Zhang Y. Nucleosome assembly is required for nuclear pore complex assembly in mouse zygotes. Nat Struct Mol Biol. 2014;21(7):609-616. doi:10.1038/nsmb.2839
[12] Oswald J, Engemann S, Lane N, et al. Active demethylation of the paternal genome in the mouse zygote. Curr Biol. 2000;10(8):475-478. doi:10.1016/s0960-9822(00)00448-6
[13] Gu TP, Guo F, Yang H, et al. The role of Tet3 DNA dioxygenase in epigenetic reprogramming by oocytes. Nature. 2011;477(7366):606-610. Published 2011 Sep 4. doi:10.1038/nature10443
[14] Amouroux R, Nashun B, Shirane K, et al. De novo DNA methylation drives 5hmC accumulation in mouse zygotes. Nat Cell Biol. 2016;18(2):225-233. doi:10.1038/ncb3296
[15] Han L, Ren C, Li L, et al. Embryonic defects induced by maternal obesity in mice derive from Stella insufficiency in oocytes [published correction appears in Nat Genet. 2018 May;50(5):768. doi: 10.1038/s41588-018-0077-0.]. Nat Genet. 2018;50(3):432-442. doi:10.1038/s41588-018-0055-6
[16] Li Y, Zhang Z, Chen J, et al. Stella safeguards the oocyte methylome by preventing de novo methylation mediated by DNMT1. Nature. 2018;564(7734):136-140. doi:10.1038/s41586-018-0751-5
[17] Au Yeung WK, Brind'Amour J, Hatano Y, et al. Histone H3K9 Methyltransferase G9a in Oocytes Is Essential for Preimplantation Development but Dispensable for CG Methylation Protection. Cell Rep. 2019;27(1):282-293.e4. doi:10.1016/j.celrep.2019.03.002
[18] Zhang Y, Xiang Y, Yin Q, et al. Dynamic epigenomic landscapes during early lineage specification in mouse embryos. Nat Genet. 2018;50(1):96-105. doi:10.1038/s41588-017-0003-x
[19] Rulands S, Lee HJ, Clark SJ, et al. Genome-Scale Oscillations in DNA Methylation during Exit from Pluripotency. Cell Syst. 2018;7(1):63-76.e12. doi:10.1016/j.cels.2018.06.012
[20] 刘中民.干细胞研究:从基础到临床[M].北京:人民卫生出版社, 2024:101-103.