
在发育过程中,多能性状态并木是稳定且一成不变的,随着发育的进行,胚胎干细胞的多能性可以分为两个不同阶段即原始态(naive state)和始发态(primed state)。在小鼠中,这两种细胞的体外状态分别为小鼠胚胎干细胞和小鼠外胚层干细胞(mouse epiblast stem cell,mEpiSC)。这两类多能态干细胞类型表现出明显不同的发育潜力,原始态多能干细胞能够构建囊胚嵌合体,而始发态干细胞则不具备类似的能力[1]。后来的研究发现在人的胚胎发育过程中同样存在这 2 种不同状态的多能性干细胞。人类原始态多能干细胞的体外稳定捕获及关键特征成为近十年来科学家们关注的重点。然而,这些具有差异发育潜力的多能干细胞背后的表观遗传差异研究尚待完善。阐明这种表观遗传差异将有助于更好地理解这些多能性状态的基本特性并且推动二者之间互相转化的研究,可为干细胞的命运决定与细胞身份维持等提供新的见解。
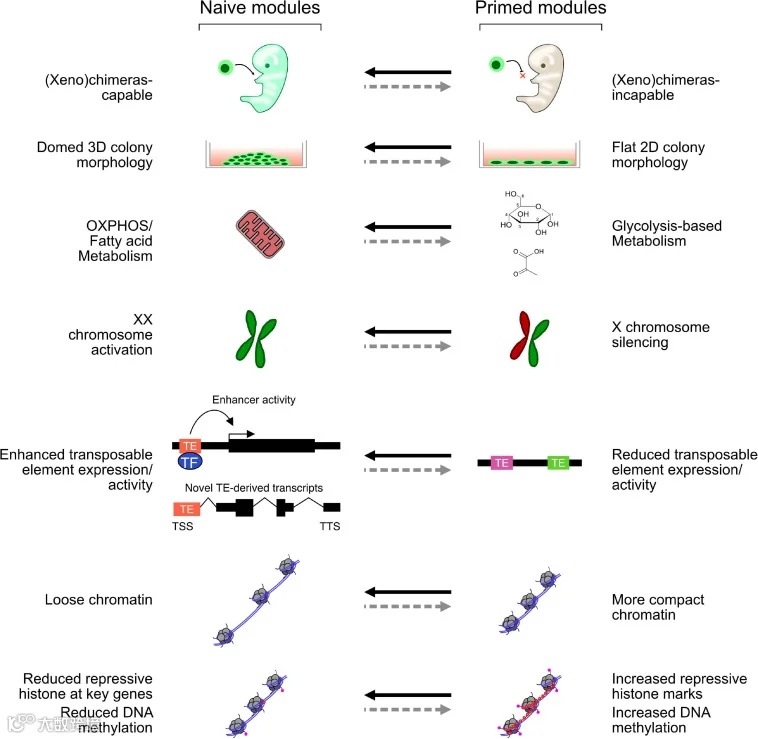
图1. 原始态和始发态多能干细胞的关键特征比较
1
组蛋白与 DNA 甲基化的差异
组蛋白修饰模式在原始态和始发态多能干细胞之间存在差异[2]。通常认为,原始态多能性的建立伴随着激活型组蛋白修饰的获得或抑制型组蛋白修饰的丢失[3,4]。在最近的研究中,人们还报道了原始态多能干细胞和始发态多能干细胞之间增强子区域的组蛋白修饰的差异。而其中即使是表达水平相近的基因的顺式调节回路也有显著差异,这说明即使在很大程度上两者共享一个通用的转录程序,它们的维持所依赖的信号传导途径也是存在差异的[5]。在小鼠胚胎干细胞中,很多基因存在多个增强子,包括显性近端增强子和休眠远端增强子(种子增强子)。但在小鼠外胚层干细胞中,原始态的显性增强子功能丢失,种子增强子承担主要的转录控制功能。种子增强子增加了序列保守性,当细胞从原始态多能性转变为体细胞调控程序时,种子增强子可确保适当的增强子利用和转录保真度[5]。一个很好的例子是 OCT4 增强子的使用,其中远端增强子优先用于原始状态,而近端增强子主要用于始发状态。这种区别意味着长距离染色质相互作用的差异,这可能有助于局部染色质 3D 结构的转变。然而,目前的研究手段很难简明扼要地统括组蛋白修饰的所有差异并精确地指明其中的关键差异要素,并且这些组蛋白差异本身是否是转录模式差异的先决条件也存在争议,有观点认为这些组蛋白修饰可能是其不同转录模式在染色体上的投射[6]。
原始态与始发态干细胞的 DNA 甲基化也存在差异。据报道,原始态小鼠胚胎干细胞的基因组 DNA 通常是低甲基化的,而始发态的小鼠外胚层干细胞却是高甲基化的[7]。在体外 2i/LIF(2 种 MEK 和 GSK3 抑制剂与白血病抑制因子 LIF)培养基中培养的原始态小鼠胚胎干细胞中表现出包括基因组印记在内的广泛的 DNA 甲基化缺失[8]。而在血清 /LIF 培养基中培养的小鼠胚胎干细胞的 DNA 甲基化水平则处于二者之间,这种层次显著的 DNA 甲基化状态差异即是小鼠胚胎干细胞和小鼠外胚层干细胞表观遗传差异的一个明显例子。
2
X染色体失活
原始态和始发态多能干细胞的另一个主要区别是 X 染色体状态的差异。X 染色体失活(X chromosome inactivation,XCI)是哺乳动物胚胎发生过程中至关重要的表观调控事件。X 染色体失活的程序相当复杂,由许多调控网络共同监管[9]。Xist-RNA 对 X 染色体的包裹被认为是 XCI 启动最早的事件之一[10]。随后 RNA po1Ⅱ和活性组蛋白修饰如 H3K4me2/3 被擦除并招募 PRC2 和PRC1,同时抑制性组蛋白标记如 H3K27me3 和 H3K9me2 被添加到失活的 X 染色体中。最近的报道中还说明转录阻抑物 SPEN 是用于启动 X 染色体失活的重要因子,可以在活性增强子和启动子处桥接 Xist-RNA 与转录机器以及核小体重构复合物和组蛋白脱乙酰酶的相互作用[11],而 XiSt RNA 结合蛋白 PTBP1、MATR3、TDP-43 和 CELF1 的组装也对 X 染色体的失活有重要的影响[12]。随后 macro-H2A 和 Ash2L 的招募被认为是相较晚发的事件。因此,确定 X 染色体失活程序启动以及随后染色体行为的分子标记值得进一步探究。
雌性 X 染色体失活状态这一表观遗传因素在拥有不同发育潜能的细胞系之间差异显著,同时在分化过程中其状态也各有异同[13]。X 染色体失活在所有体细胞谱系中往往在植入后外胚层阶段完成。有报道称,雌性小鼠外胚层干细胞中就已经出现一条失活的 X 染色体(Xi),且其富含 H3K27me3 标记,而初始态小鼠胚胎干细胞有 2 个活跃的 X 染色体(Xa)[14]。小鼠外胚层干细胞中外源性 Klf4 的强制表达也可导致 Xi 再激活[15]。由此可见,XCI状态与细胞的分化状态密切相关[16]。
参考文献
扫码关注获得更多内容

小红书@一米生物

视频号@一米生物

知乎@一米生物
公司简介


微信号|一米生物
服务热线|400-097-3606
