
在胚胎干细胞中活跃的基因在发育后期逐渐被沉默,而细胞谱系特异性基因被激活,这一过程伴随着染色质结构的改变。而胚胎干细胞在干性维持中对于自身染色质状态的调控也至关重要。多能干细胞的整体染色质结构通常是相对松散且易于转录的,在分化时染色质则被修饰成更具抑制性的状态,包括抑制性组蛋白修饰的增加、染色质可及性降低、组蛋白变体的变化以及多能性因子的去除等。
在胚胎干细胞中,组蛋白变体 H2A.Z 的含量较高。有研究证实,组蛋白变体 H2A.Z 与核小体解聚相关,这表明 H2A.Z 在调节核小体的稳定性和完整性中起关键作用,H2A.Z 可能通过调节核小体的凝集状态而参与调节 CTCF 的结合,从而调控染色质的状态和基因表达[1]。随着干细胞的分化,抑制性 H3K9me3 修饰在全基因组水平会逐渐增加,组蛋白变体 macroH2A 也会在基因组上逐渐累积,导致基因组整体水平上表现为更加致密的结构[2,3]。
维持干性的多能性因子的表达也受到染色质结构的调控。如多能性因子 NANOG 和其他几个发育调控因子的基因座是位于 DNaseI 超敏位点的位置。这些超敏位点便于多能性因子 OCT4、NANOG、ZFP281 和 Nac1 等的结合,进一步通过改变染色质高级结构来调控多能性基因的表达[4,5]。另外,染色质重塑复合物在染色质结构改变的过程中也扮演了必要角色,例如 BAF(Brg/Brahma 相关因子)或哺乳动物 SWI/SNF 复合物利用 ATP 水解产生的能量来改变染色质状态,从而控制最终影响转录组和细胞命运的转录调节子在基因组上的结合能力[6]。尽管通过 BAF 复合物影响胚胎干细胞命决定的精确分子机制仍在研究中,但是已有的研究通过对胚胎干细胞中 BAF 复合体不同亚基的敲除可以改变不同多能性转录因子的表达,并不同程度地损害了胚胎干细胞的分化能力,相应的多能性因子的表达也随之降低,这证明了 BAF 复合物在胚胎干细胞自我更新和分化中的重要作用[7]。
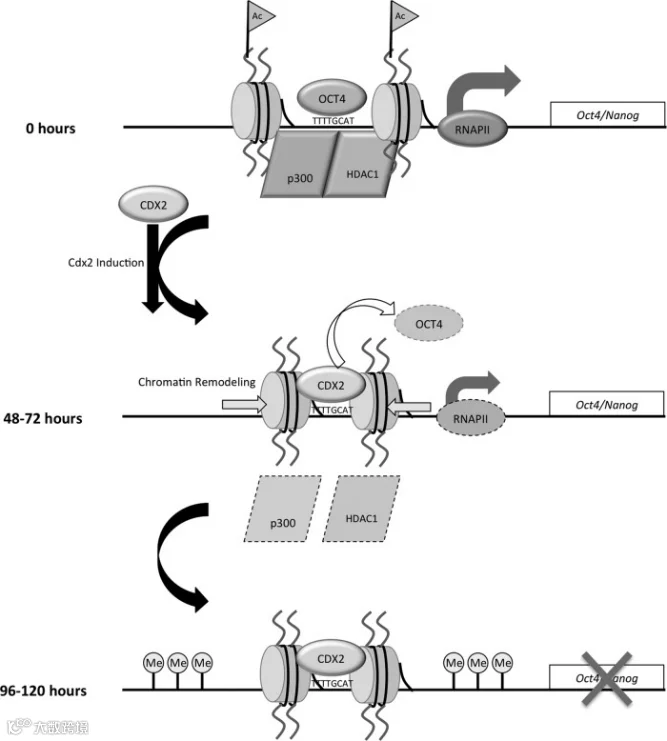
图1. 滋养层母细胞系中 Oct4 和 Nanog 的转录和染色质沉默模型
在胚胎干细胞中,miRNA 可以起到维持自我更新的作用,或者可以通过抑制多能性基因来使其适当分化[8]。有关 miRNA 调控干性的重要证据来自 miRNA 测序以及部分基因敲除的干细胞系,目前已经产生并鉴定了包括 Dgcr8 和 Dicer1等基因被敲除的几种胚胎细胞系(Dgcr8 和 Dicer1 KO ESC),这些细胞模型的详细分析均表明 miRNA 的减少和细胞干性丧失有关。Dicer 敲除的小鼠胚胎干细胞系表现出多能性的削弱出现了细胞 G1 期停滞和细胞凋亡的增加,同时细胞的自我更新也受到了阻碍[9]。然而,在人胚胎干细胞中 DICER1 的功能则不尽相同:DICER1 缺失增加了促凋亡基因的表达和细胞凋亡率,导致自我更新失败,但是并未发生与小鼠胚胎干细胞中 G1 期阻滞类似的情况[10]。miRNA 在胚胎干细胞的多能性维持以及自我更新中的作用见表 1[11]。
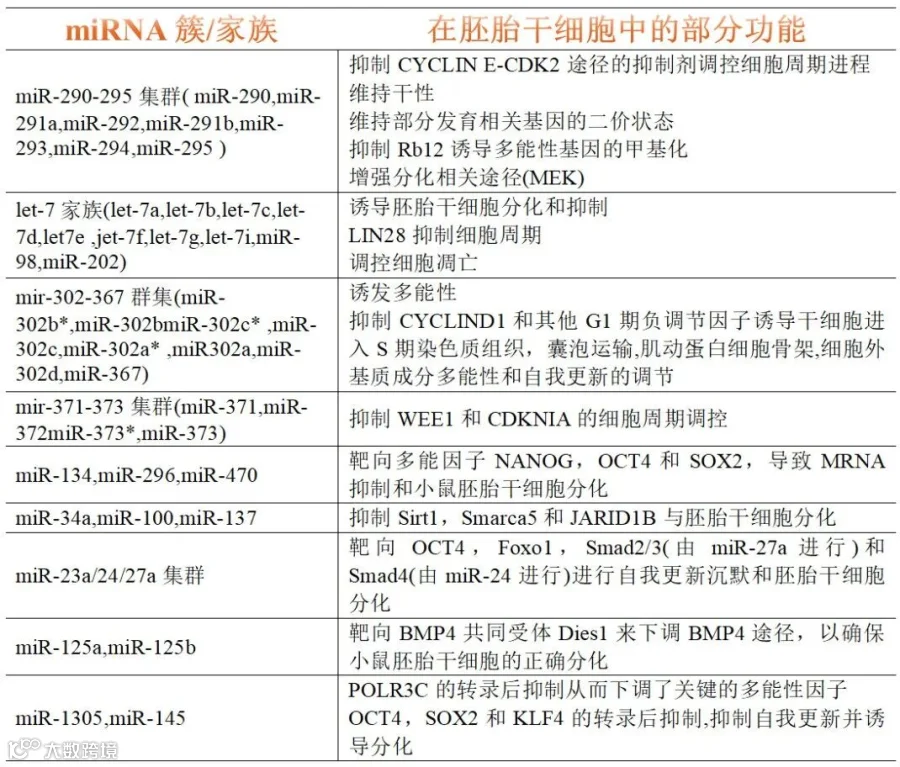
表1. miRNA 在胚胎干细胞的多能性维持以及自我更新中的作用
参考文献
扫码关注获得更多内容

小红书@一米生物

官网@一米生物

知乎@一米生物

视频号@一米生物
公司简介


微信号|一米生物
服务热线|400-097-3606
