
衰老是一种动态的细胞效应器,基因组不稳定、端粒损耗、细胞间通信改变、线粒体功能紊乱、营养缺乏、蛋白稳态丢失、干细胞衰竭、表观遗传调控改变都会促进衰老的进程[1]。
干细胞分为全能干细胞、多能干细胞、成体干细胞等,是一类具有自我更新能力及分化能力的细胞类群。终末分化细胞及成体干细胞组成了个体的大部分,机体需要成体干细胞不断的更新及分化来维持机体的稳态。成体干细胞通过再生及修复在维持个体组织结构及功能完整方面作用显著。在脑、肌肉、肠道、血液等许多组织或器官的衰老进程中,组织中的成体干细胞会出现数量变化及功能的耗竭[2]。
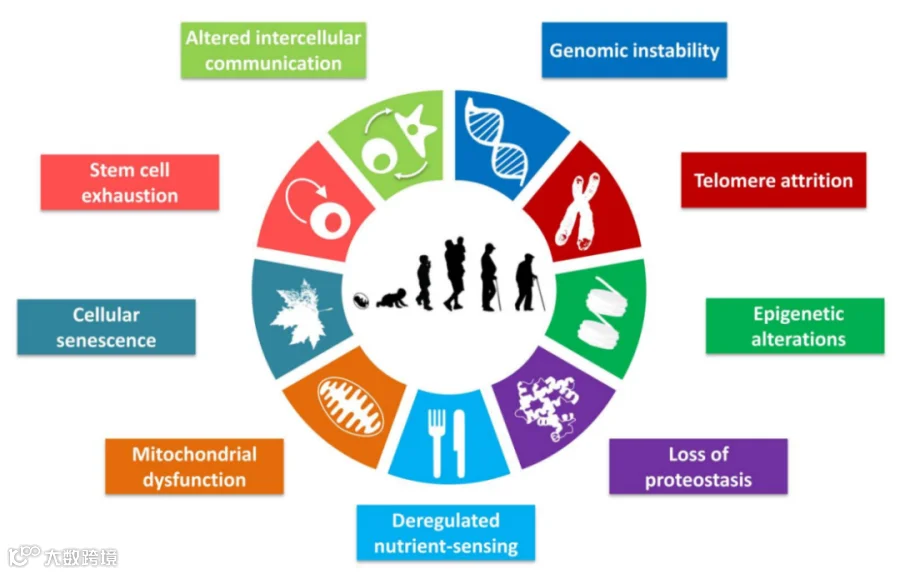
图1 衰老的相关标志
血液是哺乳动物最重要的循环系统之一,血液系统的稳态需要造血干细胞的维持。成体造血干细胞存在于哺乳动物的骨髓中,可以分化为红细胞、白细胞等髓系细胞,也可以分化为 B 细胞、T 细胞等淋巴系统细胞类型。研究表明,随着个体的衰老,哺乳动物体内的造血干细胞数目增多。同样在个体衰老过程中造血干细胞也会出现典型的衰老症状,比如归巢能力受损、移植存活能力受损、有髓系分化的偏向[3-5],而这种分化倾向容易导致骨髓恶性肿瘤的发生及免疫下降,小鼠模型对于这种分化倾向改变的可能解释是骨髓中具有分化为淋巴细胞倾向的造血干细胞在细胞数目及功能上都出现耗竭[6]。此外,小鼠中竞争性移植实验也证明衰老的造血干细胞会表现出归巢能力及移植成活能力显著减弱[7]。
神经干细胞的增殖与分化对于发育过程中中枢神经系统的形成起重要作用。而在成体脑中,神经干细胞的数目减少并且被严格限制在特定的区域。在啮齿动物中,神经干细胞分布在齿状回的粒下区和侧脑室的脑室下区[8];在哺乳动物中,实验证实了海马区存在持续的神经发生[8],成体神经干细胞仍然具有自我更新和分化为瞬时扩增祖细胞的能力[9],但这种能力随着衰老会逐渐减弱。衰老对大脑具有长久且破坏性的影响,衰老普遍会伴随认知功能下降和患脑部疾病(包括癌症和神经退行性疾病)的风险增加。一个关键的问题是,具有再生能力的细胞是否有助于大脑健康,甚至使大脑“年轻化”。与其他器官相比,大脑具有协调高级认知功能的能力,因此对机体来说至关重要。但是脑功能会随着年龄的增长而下降。同时,神经退行性疾病(如阿尔茨海默病和帕金森病),以及脑癌(如神经胶质瘤)的发病率在老年人中增[10]。尽管大脑中的所有细胞类型都会在衰老过程中受到影响,并可能导致生理性功能衰退和疾病,但是成年大脑中驻留的神经干细胞能够生成新的神经元(即神经发生)并恢复大脑部分功能。因此,在衰老过程中维持健康的干细胞储存库对于改善大脑整体健康并降低神经退行性疾病和癌症的发生率至关重要。
衰老是多种慢性和炎性疾病的主要危险因素之一,胃肠道疾病也不例外。老年人易患感染性和炎症性疾病,表现在结直肠癌发病率增高、代谢失衡、易患胃肠道感染等。与年龄相关的胃肠道变化不仅可能导致胃肠道炎症疾病的高发,还可能导致个体健康水平和寿命的下降和缩短。一些与年龄相关的并发症,如肥胖、胰岛素耐受和体弱都与胃肠道微生物群的变化有关。通常导致这类胃肠道疾病发展的条件是恶变前的化身和不典型增生。在人的胃肠道中上皮化生病变增加了胃肠道癌的风险,此类病变的特征是上皮细胞类型的异位置换。食管鳞状上皮具有胃或肠柱状上皮的特征,导致患食管腺癌的风险更高[11]。不典型增生又以细胞异常增殖和分化为特征,被认为是在上皮癌变过程中发生的,并可能促进浸润性癌的发展。在过去的十年中,果蝇和小鼠小肠干细胞(intestinal stem ce1l,ISC)的发现、鉴定以及对其年龄相关功能障碍的研究使人们认识到,这些上皮疾病可能是 ISC 活性和功能的调节改变而导致的上皮再生方向错误的结果[12]。果蝇在这方面是一种特别有效的动物模型,它可以用来详细分析细胞自身的、局部的和全身性的信号机制,这些机制在衰老动物中失调,导致胃肠道上皮内稳态的丧失。将在以下部分中详细描述该模型,并提出一种与年龄相关的果蝇胃肠道组织稳态平衡丧失的模型,该模型也会缩短动物的个体寿命。
在日常生活中,皮肤面临大量的磨损,需要不断更新和自我修复,以保持良好状态,而皮肤干细胞在皮肤细胞的更新中起了重要的作用[13]。皮肤干细胞包括位于表皮基底层的表皮干细胞,负责表皮不同层的日常再生;位于毛囊中的毛囊干细胞,确保毛囊的不断更新,再生表皮和皮脂腺;以及负责黑素细胞再生的黑素细胞干细胞。皮肤老化被定义为皮肤中某些特征的持续丧失,包括皮肤弹性和色素沉着的减少,以及皮肤干细胞的丧失,可以分为3个阶段:
①端粒磨损、DNA 损伤、基因组不稳定和氧化应激导致皮肤干细胞生长停滞、衰老或凋亡死亡;
②皮肤干细胞数量及其再生能力的逐渐下降;
③干细胞衰竭或功能障碍,加上其他有害因素,导致与年龄有关的皮肤外观或疾病的发展[14]。
在衰老过程中出现的端粒缩短、氧化应激和表观遗传的改变会造成皮肤干细胞膜、核 DNA、线粒体 DNA 的损伤,这些损伤会导致皮肤干细胞功能障碍或丧失、皮肤平衡失调和皮肤老化[15]。皮肤层通过皮肤干细胞的分裂实现伤口的愈合,在年老个体中伤口愈合形成的疤痕往往要轻于在年轻个体中形成的疤痕,通过在小鼠中的研究表明,在年轻小鼠伤口处的皮肤会高表达一种叫作基质衍生因子1(stomal cell-derived factor 1,SDF1)的分泌因子,抑制了皮肤组织再生,而在年老小鼠中通过增加 SDF1 对组蛋白甲基转移酶 EZH2 的招募抑制 SDF1 的活性,从而促进皮肤组织的再生,证明在衰老过程中组织功能并不总随着年龄增加而降低[16]。
间充质干细胞是来源于发育早期中胚层的一种多潜能细胞,具有多向分化潜能及免疫调节等功能,目前已成功建立从骨髓、脐带血、小梁骨、骨膜、滑膜、胎盘、胰腺、脂肪组织、皮肤、肺和胸腺中分离培养间充质干细胞的体系,并能够在体外诱导间充质干细胞分化为成骨细胞、软骨细胞和脂肪细胞。像其他干细胞的衰老一样,活性氧的积累、DNA 损伤、受损蛋白质的积累、端粒缩短、表观遗传学的改变都是加速间充质干细胞衰老的原因[17,18]。
参考文献
扫码关注获得更多内容

小红书@一米生物

视频号@一米生物

知乎@一米生物
公司简介


微信号|一米生物
服务热线|400-097-3606
