
在分化过程中,细胞的全能性逐渐丧失。这一过程是由交叉拮抗的转录因子的表达驱动的,转录因子促进向单一的细胞命运发展,同时抑制其他的分化途径[1]。除此之外,细胞命运的决定通过 DNA 和染色质水平上逐步获得相应的表观遗传修饰来加强[2,3]。由于发育过程中获得的关键表观遗传修饰是稳定的,并通过随后的细胞分裂进行遗传,因此建立了一种“表观遗传记忆”,作为分化细胞状态表型稳定性的基础,而这种表观遗传记忆的建立以及上游细胞类型的表观遗传记忆的去除则在细胞命运决定的过程中尤为重要[4]。
虽然在生物的正常发育过程中,细胞命运的决定往往是程序且稳定的,但在体外培养的细胞系内,细胞命运的决定可以被人为调控甚至逆转。成功的重编程过程实际上需要完全擦除现有细胞的表观遗传记忆,然后建立新的特定细胞类型的表观遗传标记。因此,尽管仅通过关键转录因子的异位表达就可以实现细胞身份的改变,但转化效率仍然很低,其部分原因与现有的染色质修饰构成的重编程障碍有关[5]。而随着各类显微操作技术与细胞培养技术的革新,人们开始寻找与表观遗传记忆的消除和多能性或全能性重建有关的机制。下文也将围绕着细胞重编程过程中细胞命运的决定机制展开简要讨论。
诱导性多能干细胞的表观遗传调控
随着细胞培养技术的进步,人们可以通过异位表达所需细胞类型的先驱转录因子来实现细胞命运的转变。而近年来最为轰动的案例为诱导多能干细胞。通过 4 种多能性相关转录因子:OCT4,SOX2,KLF4 和 MYC(OSKM)的异位表达可将体细胞直接重编程为诱导多能干细胞。这些转录因子的表达破坏了分化的体细胞的转录调控网络,并导致建立拥有胚胎干细胞样表型的体外培养细胞系[6]。除了改变转录网络,在诱导多能干细胞重编程过程中,OSKM 转录因子的过表达已被证明可诱导大规模染色质变化,最终导致建立与胚胎干细胞高度相似的染色质结构[7,8]。尽管诱导多能干细胞具有类似于胚胎干细胞的许多特征,但其原始体细胞时期的表观遗传记忆仍可能保留在其基因组中,而体细胞记忆可能会损害整个诱导过程,这与所得诱导多能干细胞系的质量也息息相关。通常,转录因子 OSKM 可通过结合其他转录因子并直接募集各种组蛋白修饰因子来重塑其结合区域的染色质结构[9]。转录因子的结合也可诱导基因座特异的 DNA 去甲基化发生[10]。另一方面,越来越多的证据表明,染色质在重编程的早期阶段就开始发挥重要的作用。在诱导多能干细胞重编程过程中,OSKM 的初始参与受到抑制性组蛋白修饰的阻碍[11],而其后续过程中也被证明存在多种表观遗传因子的阻碍[12,13]。根据上述的研究与讨论,有效的重编程不仅需要允许快速有效地结合引入的转录因子,而且能够促进染色体组分的转换,从而能够快速有效地清除预先存在的 DNA 和组蛋白修饰并重建[14]。
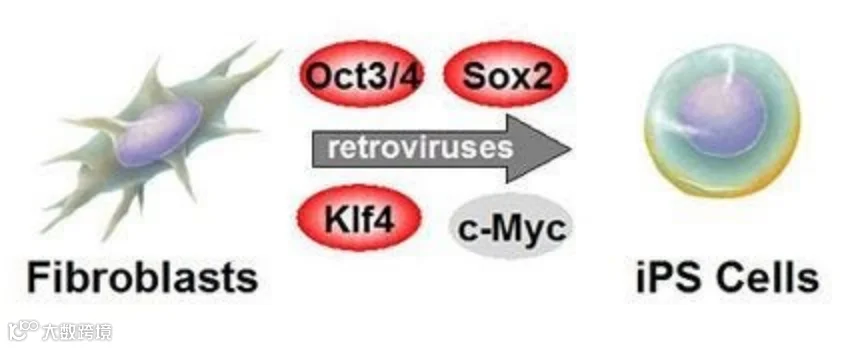
图1 体细胞构建iPSC
参考文献
扫码关注获得更多内容

小红书@一米生物

视频号@一米生物

知乎@一米生物
公司简介


微信号|一米生物
服务热线|400-097-3606
