
遗传调控及表观遗传调控通过端粒、基因突变或异位、拷贝数差异、DNA 甲基化、组蛋白修饰、染色质重塑、长链非编码 RNA 等方式改变对成体干细胞进行调控,这些成体干细胞包括神经干细胞、间充质干细胞、小肠干细胞、皮肤干细胞、造血干细胞等[1-6],从而对衰老起到调控作用。
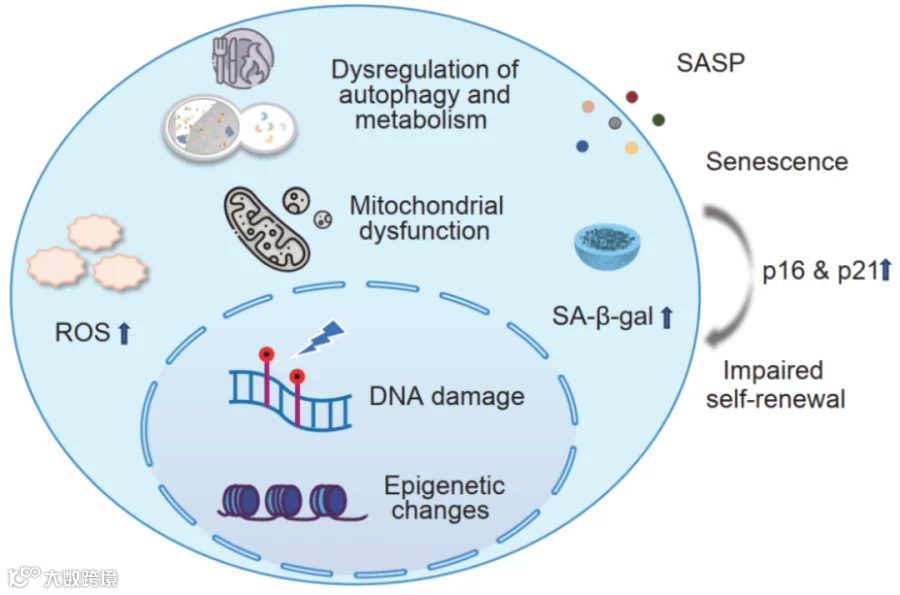
图1 细胞衰老的机制
基因的表达是一个随机的过程,这种随机表达的过程是通过染色质拓扑结构或者染色体修饰的改变实现的。染色体由串珠状的核小体组成,紧缩的核小体结构对于基因表达呈抑制作用。染色质重塑是指在相应的复合物的作用下,染色质的包装状态、核小体中组蛋白以及对应 DNA 分子会发生改变。这些复合物主要包括四个家族:SWI/SNF、ISWI、NuRD 和 INO80,这些复合物都具有 ATP 酶活性[7],能够切割组蛋白与 DNA 之间的联系,引起核小体的重新分布。
此外染色质重塑复合物的作用一般是与组蛋白修饰及组蛋白伴侣协同作用,以完成核小体包装、驱逐及滑动等精确有序的调节过程。目前已有许多研究表明,癌症发生过程中伴随着染色质重塑调节蛋白功能缺失。越来越多的研究表明,染色质重塑调节蛋白的功能与衰老的进程密切相关。生理性衰老及早衰成体干细胞模型中都出现大范围的染色质结构的改变,同时伴随着 DNA 损伤增加。
儿童早衰症患者中,NuRD 蛋白家族复合物 RBBP4 和 RBBP7 表达下调。同样在正常细胞系中进行 RBBP4 和 RBBP7 的蛋白敲除,发现缺陷的细胞表现出早衰的表型。另外生理性衰老的细胞也表现出 NURD 蛋白家族复合物成分的降低,虽然这种蛋白表达降低的具体分子机制还未发现,但是这些研究都表明染色质重塑蛋白对衰老的进程有重要的调控作用。Isw2 蛋白复合物的缺失会导致酵母寿命的延长,这种延长作用是通过激活 RAD51 基因的表达而实现的,RAD51 可以调控同源重组方式修复 DNA 的效率,从而实现寿命延长。SWI/SNF 复合物催化亚基 Chd1 的缺失同样也可以延长酵母的寿命。INO80D 复合物亚基 Ser818Cys 突变会促进人类主动脉的衰老。此外线虫中研究表明,SWI/SNF 核心催化亚基的失活会导致 DAF-16/FOXO 延长寿命效果丢失。在人类中发现,SWI/SNF 复合物中 BRG1 在先天性心脏病患者的心肌层中表达明显下调。BRG1 活化并与 G9a/GLP 和 DNMT3 组成复合物,结合到 MYH6 的启动子区域,通过对其启动子区域进行组蛋白甲基化或 DNA 甲基化,以抑制 MYH6 的表达,造成人及小鼠的心肌肥大[8]。
总体来说,染色质的状态受到染色质重塑复合物的调控。染色质状态的改变可能会影响衰老相关基因的表达。从酵母、线虫、小鼠到人类,这些染色质调控蛋白结构及功能都是保守的,进一步探究染色质重塑蛋白调节干细胞衰老的机制及如何通过干预这个过程来实现健康衰老甚至逆转衰老需要进一步研究[9]。
参考文献
扫码关注获得更多内容

小红书@一米生物

视频号@一米生物

知乎@一米生物
公司简介


微信号|一米生物
服务热线|400-097-3606
