

类器官在体外再现了器官生物学的原理,并提供了简化的、易于获得的“最小系统”,用于辨别不同组织成分对复杂形态发生过程的相对贡献。事实上,类器官已经揭示了组织在稳态、再生和发育中的机制。由于邻近组织的潜在混杂影响,这些组织自主机制在体内很难识别。例如,在 ASC 衍生的肠类器官中,从隐窝分离的干细胞在缺乏上皮下间质室的情况下重建肠上皮[1];在多能干细胞衍生的视杯类器官中,视泡自主地进行形态发生,而不需要来自上覆表面外胚层的信号[2]。
类器官的一个关键特征是它们比哺乳动物模型更易于实验,这有助于更深入地理解器官发生和成人器官生物学。表达 Lgr5 基因的小鼠肠道干细胞(intestinal stem cells,ISC)形成自我更新的克隆的能力已被证明依赖于帕内特细胞(Panethcell)[3]。帕内特细胞表达高水平的 Wnt 配体,Wnt 信号转导对 ISC 的维持至关重要[4]。一项研究使用了表达标记 Wnt 亚型的细胞来探索 Wnt 梯度在肠道生物学中的作用[5]。在研究中肠道类器官的使用提供了一个实验性的系统来产生高分辨率的信息[5]。这项研究表明,Wnt 蛋白不会扩散,而是从帕内特细胞转移到相邻的 ISC,并通过 ISC 分裂和子细胞中膜结合的 Wnt 稀释,沿隐窝形成 Wnt 梯度。这一结果表明帕内特细胞作为 ISC 自我更新的局部 Wnt 信号源[5],并支持一种模型,即帕内特细胞在隐窝底部的空间限制定义了 ISC 自我更新和分化之间的物理分离。这种物理分离早前在体内对 ISC 进行克隆分析实验时就已被证实,这表明对隐窝生态位的竞争决定了 ISC 是自我更新还是分化[6]。
在视杯类器官上进行的实验,提供了对视杯形态发生机制的见解,这一过程涉及视网膜色素上皮(retinal pigmented epithelium,RPE)、中视网膜神经(neural retina,NR)的内陷。用抑制细胞骨架活性收缩或细胞增殖的药物治疗小鼠视泡向视杯转化的不同阶段,表明视杯的形态形成是一个多阶段的过程。囊泡内陷的过程始于细胞骨架重组引起的机械特性的领域特异性变化。这些变化导致了收缩性的降低,从而使 NR 的硬度降低,并维持了 RPE 的高水平收缩性和硬度。在随后的过程中,细胞增殖的作用成为主导作用,产生必要的推力,以使 RPE[2] 内较不坚硬的 NR 内陷。
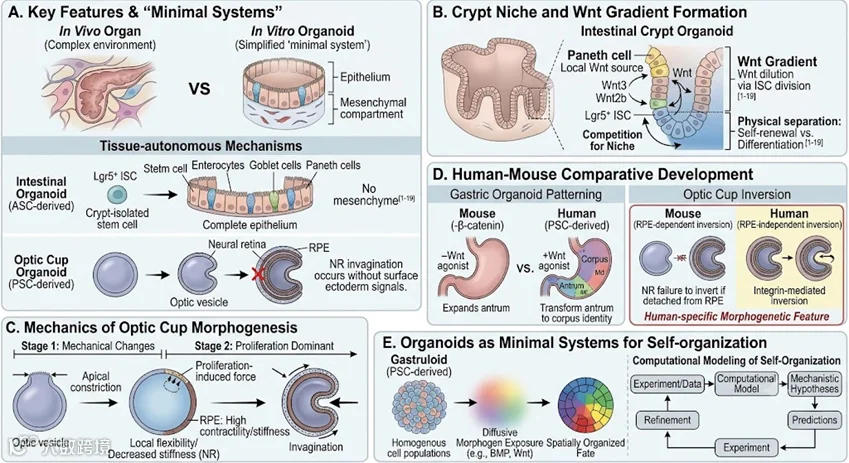
图1 类器官的应用——稳态、代谢和发育
由于人类胚胎和胎儿组织的可得性极其有限,以及围绕其使用的伦理问题,人类发育和器官发生的研究受到了限制,因此从人类干细胞衍生的类器官中获益良多。此外,将人类器官与动物摸型或从动物模型衍生出来的类器官进行比较,可以突出人类与其他物种在发育方面的相似性或差异性。例如,小鼠胚胎缺乏编码 Wnt 通路核心转录因子的 β-联蛋白基因,表现出胃底的身份缺失和胃窦结构域扩张[7]。这一结果表明Wnt 信号在指定基底身份方面起着重要作用。为了确定这是否也适用于人类胃模式,在前肠的后阶段,将人多能干细胞衍生的胃类器官[8]暴露于 Wnt 激动剂中,区域特异性标记的表达分析证实了从胃窦到胃底身份的转换[7]。
胃类器官强调了人类和小鼠的共同发育机制,而视杯类器官强调了人类特有的发育特征[2,9]。事实上,人类视杯类器官比老鼠的视杯类器官要花更长的时间才能达到相应的发育阶段,它们也比小鼠类器官更大,神经质更厚。这些物种特异性差异被人类和小鼠胚胎组织的分析证实了[10]。此外,视杯类器官揭示了物种特异性的形态发生机制。正常情况下,在从视神经囊到视杯的发展过程中,神经质由顶端的凹状结构向顶端的凸状结构转变。在小鼠类器官中,这种所谓的外翻需要神经质和 RPE 之间的连续性。在囊泡期从 RPE 分离的小鼠 NR 不会发生外翻[2,11]。相比之下,在囊泡期从 RPE 分离的人 NR 仍然能够通过整合素介导的肌球蛋白独立机制自主地发生外翻[12]。
类器官也可以作为研究自组织原理的最小系统。一个有趣的例子是对原肠胚形成的胚胎类器官模型的研究[13-18]。在这些系统中,细胞由于均匀暴露于形态发生素而采取空间组织的命运,这意味着它们是自组织的[14,16-19]。在其中一些研究中,计算机数学模型和实验数据的组合已被用于测试特定分子机制概括自组织结局的能力[14,17]。然后利用这些模型成功地预测了原肠胚在不同启动条件下的行为[14,17]。一般来说,数学模型有助于阐明自我组织的机制,预测不同条件对结果的影响,从而提高我们在体外控制复杂多细胞行为的能力[19]。
参考来源
扫码关注获得更多内容

小红书@一米生物

视频号@一米生物

知乎@一米生物
公司简介


微信号|一米生物
服务热线|400-097-3606
