
···点击左上方蓝字 · 关注上海逐典···
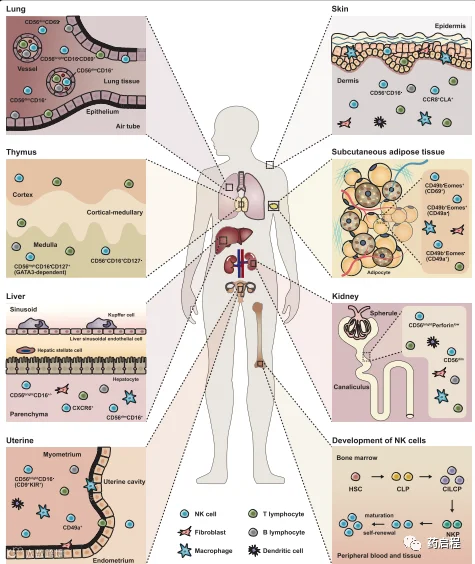
图1 NK细胞的发育和亚群
NK细胞是先天免疫系统的重要参与者,构成抵御癌细胞的第一道防线。自然杀伤(NK)细胞由Herberman在1976年定义,一种新的淋巴细胞群。NK细胞遍布全身,占血液和器官中所有淋巴细胞的5-20%,由于存在独特的趋化因子受体,NK细胞分布在健康组织中有所不同,在骨髓、脾脏、肝脏、肺、皮肤、肾脏、子宫和次级淋巴组织中浓度较高。NK细胞源自CD34+共同淋巴祖细胞,并在骨髓(BM)中分化为未成熟和成熟的NK细胞。然后分布到淋巴和非淋巴外周器官和组织,包括PB、脾、肺、肝和子宫。
NK细胞的发育过程
NK细胞具有类似于CD8+ T细胞在适应性免疫中发挥作用但缺乏CD3和T细胞受体(TCR)的细胞毒性能力。NK细胞主要在血液中循环,约占外周血单个核细胞(PBMC)的5-10%,存在于骨髓和脾脏等淋巴组织中。与其他ILC类似,NK细胞起源于骨髓中的共同淋巴祖(CLP)细胞(图 1),平均更新周期约为2周。
在开发过程中,一个称为“教育”的过程描述了表达免疫受体酪氨酸抑制基序(ITIM)的NK细胞与主要组织相容性复合体-I (MHC-I) 的相互作用,帮助NK细胞获得许可并避免攻击健康正常细胞。有趣的是,肿瘤细胞总是缺乏或仅表达低水平的MHC-I以逃避CD8+ T细胞介导的细胞毒性,而获得许可的NK细胞则被完全激活。然而,肿瘤细胞也表达激活NK细胞的分子,例如MHC I类多肽相关序列A (MICA)和MICB,支持使用NK细胞作为抗癌剂。此外,未经许可的NK细胞在体内也发挥着重要作用,例如消除鼠巨细胞病毒 (MCMV)感染和MHC-I+细胞。
迄今为止,NK 细胞的存活和发育被认为主要依赖细胞因子(尤其是 IL-2 和 IL-15)和转录因子(Nfil3、Id2和Tox用于发育,EOMES和T-bet用于成熟)。GRB2相关结合蛋白 3(GAB3)对IL-2和IL-15介导是必不可少的,其缺乏会导致 NK 细胞扩增受损。此外,靶向相关信号是促进NK细胞诱导的癌症细胞毒性的潜在选择。如前所述,细胞因子诱导型含SH2蛋白(CIS)的消融可负向调节IL-15以限制NK细胞功能,可预防转移并增强CTLA-4和PD-1体内阻断治疗。
NK细胞的表面分子
由于NK细胞表面标志物的可变表达,很难用一两个简单分子或传统的免疫组化来准确识别这种细胞类型,更重要的是,它们的功能状态。然而,在人类的临床和研究环境中,CD3-CD56+细胞通常被认为是NK细胞,可以进一步分为CD56bright和CD56dim亚群。CD56不仅是一个标志物,而且在NK细胞的终末分化中也起着重要作用,因为它被单克隆抗体阻断明显抑制了从CD56bright到CD56dim的转变,从而限制了细胞毒能力。
一致地,CD3-NK1.1+和CD3-CD49b+细胞被定义为小鼠的NK细胞。在最近的研究中,基于将更多功能性蛋白质而不是表面分子添加到分类中的共识,提出了属于天然细胞毒性受体(NCR)的天然细胞毒性受体46(NKp46) 也应包含在该面板中的概念NK细胞系统。
NK细胞中的激活和抑制信号
作为先天免疫中的主要效应细胞类型,NK细胞能够在很早的阶段杀死肿瘤细胞和病毒感染的细胞。由于缺乏大量产生的受体来特异性区分体内无法计算的抗原,它们依靠“缺失自我”和“诱导自我”模式通过维持激活共刺激和抑制之间的精确平衡来识别靶细胞。信号(主要通过功能性受体)。这些相互作用的信号最终决定了NK细胞的激活和功能状态。
激活信号包括细胞因子结合受体、整合素、杀伤受体(CD16、NKp40、NKp30和NKp44)、识别非自身抗原的受体(Ly49H)和其他受体(例如,NKp80、SLAM、CD18、CD2 和 TLR3/9)。总的来说,NK细胞的激活受体根据各自的配体可分为至少三种类型,包括MHC-I特异性受体、MHC-I相关受体和MHC-I非相关受体(表1)。
需要强调的是,属于第三组的NCRs包括三个分子(NKp30、NKp44和NKp46),并且NKp30被证明能够识别肿瘤细胞上表达的B7-H6,可以作为一种新的治疗方法未来的选择。
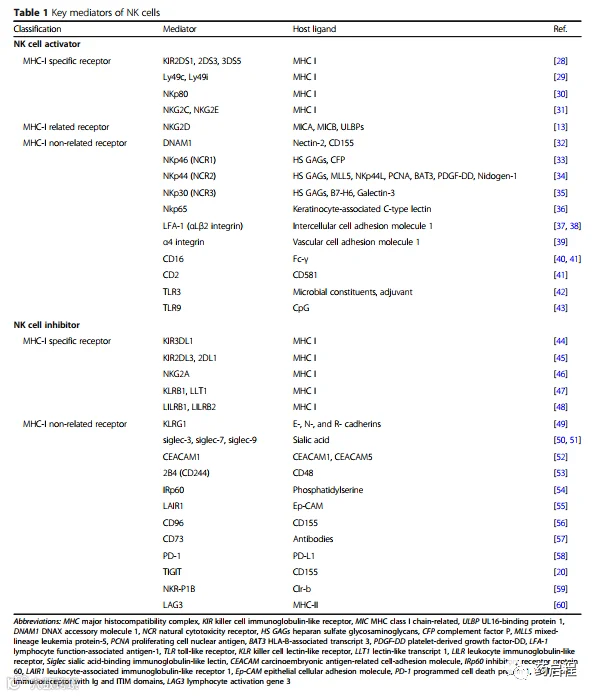
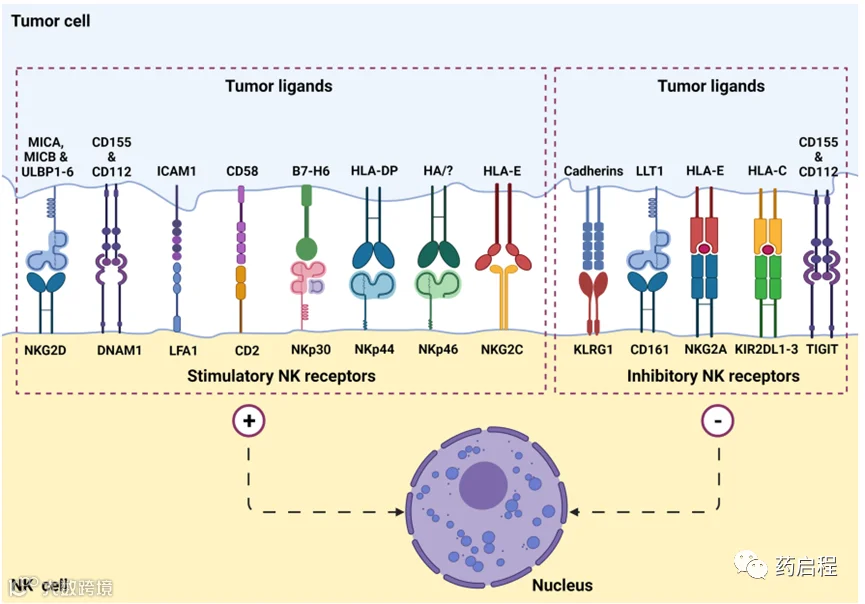
图2 NK细胞表面受体和肿瘤细胞上的配体参与肿瘤识别。NK细胞表达一组刺激(或激活)受体和抑制性受体,通过MHC-1受体外观识别健康细胞和异常细胞,如病毒感染或潜在的致瘤细胞。
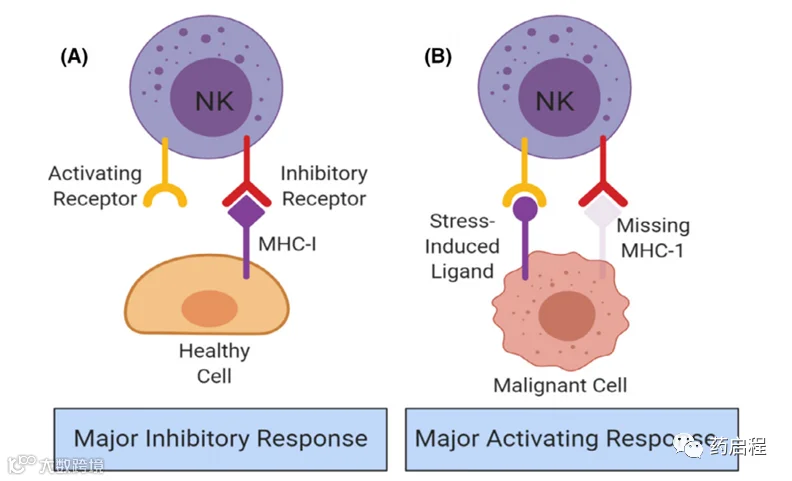
图 3. 根据“失踪的自我”和“改变的自我”模型对自然杀伤 (NK) 细胞反应的调节。((A) 作为抑制性NK细胞受体配体的主要组织相容性复合体 (MHC)-I 的存在,以及健康细胞表面缺乏应激诱导的配体,导致NK细胞的主要抑制信号 .(B) 激活NK细胞受体的应激诱导配体的存在和肿瘤细胞对MHC-I的下调导致NK细胞的主要激活信号。[色图可在 wileyonlinelibrary.com 查看]
抑制信号主要包括识别MHC-I的受体,如Ly49s、NKG2A和LLT1,以及一些与MHC-I无关的受体(表 1)。此外,MHC-I特异性抑制性受体根据结构和功能一般可分为三种类型:杀伤细胞免疫球蛋白样受体(KIRs)、杀伤性凝集素样受体(KLRs)和白细胞免疫球蛋白样受体(LILR)。
根据成熟部位的NK细胞亚群
常规NK(cNK)细胞主要存在于外周血中,并迁移到特定位置发挥作用。NK细胞还包括组织驻留NK(trNK)细胞。NK细胞分化的复杂过程发生在几个不同的组织中,包括骨髓、肝脏、胸腺、脾脏和淋巴结,并且可能涉及这些组织之间不同成熟阶段的细胞循环。在骨髓、血液、脾脏和肺中,NK细胞完全分化,而在淋巴结和肠中的NK细胞是未成熟和前驱的。骨髓和血液NK细胞的单细胞转录组分析有助于说明其在发育过程中的特征变化。例如,TIM-3、CX3CR1和ZEB2的高表达代表更成熟的状态。
总之,各种组织中的NK细胞具有不同的特征,具有不同的功能并与其他基质细胞形成密切的关系(图1)。在肺中,trNK细胞显示出与循环NK 细胞(主要是CD56dim)不同的表型,并被认为表达不同水平的CD16、CD49a和CD69,其中CD56dimCD16+细胞代表整个NK家族的大部分。值得注意的是,CD69+细胞是CD56brightCD16-NK细胞的主要类型。然而,在胸腺中,大多数NK细胞是 CD56highCD16-CD127+,与CD56+CD16+亚组相比,高度依赖GATA3。此外,它们产生更多的效应分子,包括TNF-α和IFN-γ。
类似地,肝trNK细胞可分为两组,包括CD56brightCD16+/-和CD56dimCD16+,均缺乏CD3和CD19。此外,CD49a+CD56+CD3-CD19-NK细胞已在肝活检中被鉴定。此外,肝NK细胞可以对结构多样的抗原产生记忆,这取决于表面分子CXCR6。在子宫中,大多数NK细胞是CD56brightCD16-,表达高水平的KIR。对于皮肤NK细胞,有趣的是只能检测到很少的 CD56+CD16+,这在外周血中很常见。有趣的是,trNK细胞在皮下(CD56dim)和内脏(CD56bright)脂肪组织之间是不同的,一般可以根据CD49b和Eomes分为三组,显示不同的CD49a(CD49b+ Eomes-亚组)和CD69表达水平(CD49b-Eomes+ 亚组)。
根据功能分子划分的NK细胞亚群
根据表面CD56的表达,NK细胞可分为CD56bright和CD56dim。CD56dim NK细胞主要存在于外周血中,并且总是CD16阳性,表达高水平的KIR和LFA-1,并显示出细胞杀伤能力。
CD16是介导抗体依赖性细胞毒性 (ADCC) 的关键受体,可诱导免疫受体酪氨酸激活基序 (ITAM) 磷酸化。根据时间分辨单细胞试验,NK细胞的细胞毒性通过坏死和凋亡被抑制。因此,FasL/FasR 相互作用、forin/颗粒酶释放和 Ca2+流入对NK细胞功能都很重要。然而,CD56bright NK细胞与辅助细胞相似,主要分泌IFN-γ、TNF-β和GM-CSF等细胞因子。研究人员甚至进一步将这些细胞分为NK1和NK2类别,与 Th1和Th2 一致,主要分别分泌IFN-γ和IL-5。
除了已建立的细胞毒性cNK细胞外,已经证明NK细胞可以分化为抗原呈递NK(AP-NK)细胞、辅助NK(NKh)细胞和调节性NK(NKreg)细胞,每个细胞定义为 表面分子和个体功能。一种具有专业APC能力的新CD8αα+ MHC-II+表型被认为代表了不寻常的AP-NK细胞,识别和消除自身反应性T细胞并最终像cNK细胞一样杀死它们。被预防性疫苗 FSME激活的人浆细胞样树突状细胞(DC)上调了其表面CD56的表达。
不变的自然杀伤T细胞 (iNKT) 构成了一个表达NK细胞标志物的T细胞亚组。NKT被CD1d呈递抗原激活,不仅可以分泌Th1型细胞因子,还可以分泌Th2型细胞因子参与免疫。Th1极化的iNKT细胞表现出肿瘤耗竭表型,而Th2极化的iNKT细胞与极化的T细胞相似,有助于肿瘤进展。最近的研究还强调了iNKT细胞的新功能亚型。然而,近年来,由于它们与先天免疫的密切关系,iNKT细胞有可能被定义为ILC的一个特殊亚群。
从NK细胞的发展和功能可见,它们在癌症生物学中起到不同作用。NK细胞通过与癌细胞、基质细胞、细胞外基质,尤其是代谢产物的相互作用,发挥抗肿瘤免疫作用。
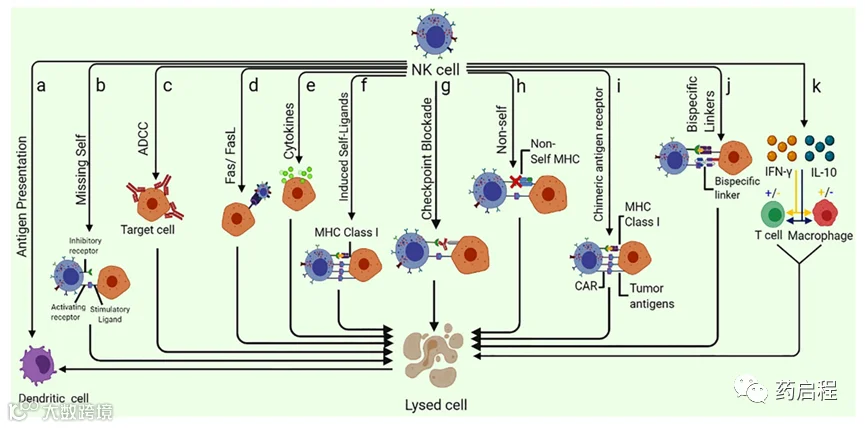
图 4 NK细胞介导的肿瘤杀伤和免疫系统调节的不同方式:(A)NK细胞能够通过杀死未成熟的DC,同时促进 IFN-γ和TNF-α介导的DC成熟来增强向T细胞的抗原呈递.(B)NK细胞可以特异性识别缺乏自身MHC I类分子(Missing-self)表达的细胞。(C)ADCC可以杀死靶细胞。(D)Fas/FasL通路是一种非常有效的NK细胞介导的细胞杀伤(E)细胞因子途径可以发挥抗肿瘤潜力,因为细胞因子(例如NK细胞)会分泌多种细胞因子,例如 TNF-α。(F)NK细胞受体NKG2D能够识别“诱导自我”配体,这些配体响应于肿瘤相关通路的激活以非常高的速度表达。(G)检查点阻断可以通过阻止N 细胞抑制受体与其配体的相互作用来抑制NK细胞抑制。(H)由于过继NK细胞转移,供体和受体之间的错配,抑制性KIR,NK细胞消除了缺乏自身MHC的同种异体肿瘤细胞。(I)专门针对过表达的肿瘤抗原设计的CAR-NK细胞也可用于消除特定的肿瘤细胞。(J)专门设计的双特异性分子也被用于特异性消除肿瘤细胞,因为这些特殊分子结合激活一侧的 NK 细胞受体和另一侧的肿瘤抗原。(K)NK细胞可以通过IFN-γ 和IL-10的产生增强或减弱巨噬细胞和T细胞的活性。
来源:药启程
* 推文用于传递知识,如有版权等疑问,请联系我们。
参考文献(可上下滑动)
1. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–74.
2. Wu SY, Fu T, Jiang YZ and Shao ZM. Natural killer cells in cancer biology and
therapy. Molecular Cancer (2020) 19:120-146.
3. Netea MG, Joosten LA, Latz E, Mills KH, Natoli G, Stunnenberg HG, O'Neill
LA, Xavier RJ. Trained immunity: A program of innate immune memory in
health and disease. Science. 2016;352:aaf1098.
4. Xie GZ, Dong H, Liang Y et al. CAR-NK cells: A promising cellular immunotherapy for cancer. EBioMedicine 59 (2020) ;
5.Gubin MM, Zhang X, Schuster H, Caron E, Ward JP, Noguchi T, Ivanova Y,
Hundal J, Arthur CD, Krebber WJ, et al. Checkpoint blockade cancer
immunotherapy targets tumour-specific mutant antigens. Nature. 2014;515:
577–81.
6. Pitt JM, Vetizou M, Daillere R, Roberti MP, Yamazaki T, Routy B, Lepage P,
Boneca IG, Chamaillard M, Kroemer G, Zitvogel L. Resistance Mechanisms to
Immune-Checkpoint Blockade in Cancer: Tumor-Intrinsic and -Extrinsic
Factors. Immunity. 2016;44:1255–69.
7. Herberman RB, Holden HT, Ting CC, Lavrin DL, Kirchner H. Cell-mediated
immunity to leukemia virus- and tumor-associated antigens in mice. Cancer
Res. 1976;36:615–21.
8. Chiossone L, Dumas PY, Vienne M, Vivier E. Natural killer cells and other
innate lymphoid cells in cancer. Nat Rev Immunol. 2018;18:671–88.
往期精彩:



