
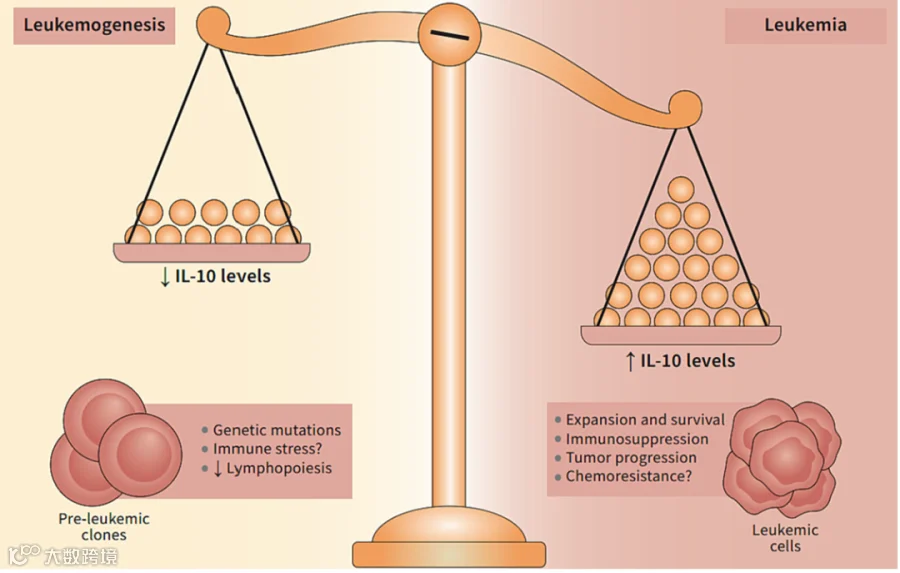
白细胞介素 - 10(IL-10)在多种生理和病理过程中发挥多样化作用。
一、IL-10 的发现与基本特性
- 发现历程
30 年前作为 Th2 细胞分泌的细胞因子合成抑制因子被发现,可抑制 Th1 细胞的细胞因子产生,其基因与 EB 病毒编码的 BCRF1 蛋白高度同源,提示病毒可能利用其免疫抑制特性。 - 家族成员
IL-10 是 IL-10 家族的创始成员,该家族还包括 IL-19、IL-20、IL-22 等 9 种细胞因子。 - 生物学作用
作为关键抗炎介质,在宿主抵御病原体和微生物群过度反应、无菌伤口愈合、自身免疫、癌症和体内平衡中发挥重要作用。
二、IL-10 的产生细胞与调控机制
(一)产生细胞
- 免疫细胞
Th2 细胞、CD4/CD8 T 细胞、B 细胞、巨噬细胞、DC、中性粒细胞、肥大细胞等。 - 非免疫细胞
上皮细胞、肿瘤细胞、 resident macrophages(如小胶质细胞、心脏巨噬细胞)。 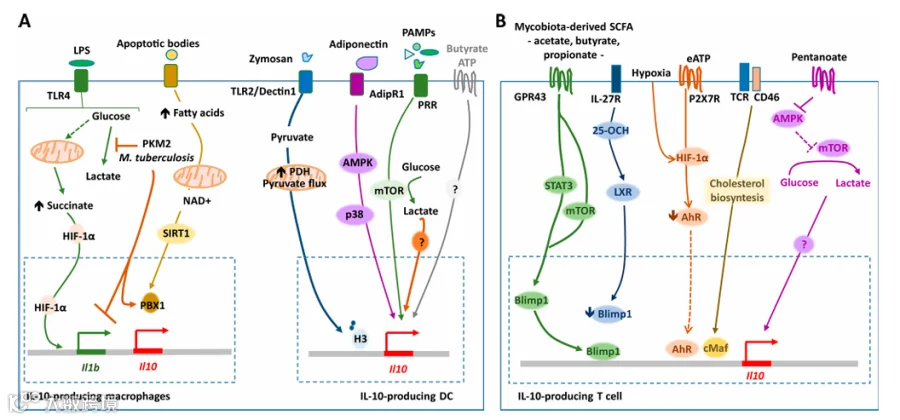
10.1084/jem.20190418
(二)调控机制
- 信号通路
- MAPK 通路
ERK1/2 和 p38 激活后通过 CREB、ATF1 等转录因子促进 IL-10 表达。 - NF-κB 通路
其家族成员结合 IL-10 基因位点促进表达。 - PI3K/Akt 通路
通过抑制 GSK3β 和激活 mTOR 促进 IL-10 产生。 - 转录因子
- cMaf
作为髓系和 T 细胞 IL-10 产生的 master regulator,与 Blimp-1、AhR 等协同作用。 - Blimp-1
对 Th1 细胞和 CD8 T 细胞的 IL-10 表达至关重要。 - STAT3
在 IL-10 信号下游发挥关键作用。 - 代谢调节
- 糖代谢
PKM2 抑制糖酵解重编程,促进 IL-10 产生;M. tuberculosis 感染抑制糖酵解可增加 IL-10。 - 脂代谢
短链脂肪酸(如戊酸)通过代谢 - 表观遗传重塑诱导 IL-10。 - 核苷酸代谢
抑制 NAMPT 酶可促进单核 / 巨噬细胞向 IL-10 极化。 - 表观遗传
染色质重塑(如组蛋白乙酰化)和非编码 RNA(如 miR-155)参与 IL-10 调控。
三、IL-10 的信号传导途径
- 受体组成
由高亲和力的 IL-10Rα 和泛表达的 IL-10Rβ 组成异二聚体。 - 下游信号
JAK1 和 TYK2 激酶激活后磷酸化 STAT3,使其入核调控抗炎基因表达;同时激活 PI3K/Akt/mTOR 和 AMPK 等通路。 - 信号特异性
STAT3 可被 IL-6 等促炎因子激活,但 IL-10 通过 SOCS3 等机制避免与促炎信号交叉。
四、IL-10 的生物学功能
(一)经典抗炎作用
- 髓系细胞抑制
减少巨噬细胞、DC 的促炎因子(如 TNF-α、IL-1β)和 MHCII、共刺激分子表达,抑制中性粒细胞的细胞因子产生。 - T 细胞调节
通过 APC 间接抑制 T 细胞应答,或直接诱导 CD4 T 细胞无反应性,同时增强 CD8 T 细胞的增殖和细胞毒性。
(二)非经典 roles
| 作用对象 | 具体功能 | 机制 |
|---|---|---|
| 神经元 |
|
|
| 脂肪细胞 |
|
|
| 上皮细胞 |
|
|
| CD8 T 细胞 |
|
|
- 基于 IL-10 的肿瘤治疗策略
- PEG 化修饰
PEG 化是将聚乙二醇(PEG)连接到 IL-10 上,可增强 IL-10 的稳定性,延长其循环时间,降低免疫原性,减少肾脏清除,降低全身给药剂量,进而减轻剂量限制毒性。PEG 化的 IL-10(如 pegilodecakin)在早期临床试验中对部分晚期实体瘤患者(如葡萄膜黑色素瘤、晚期肾细胞癌患者)展现出一定的抗肿瘤活性,能提升血清 IL-10 浓度,且不良反应多为轻中度。 - 融合蛋白技术
IL-10/Fc 融合蛋白将 IL-10 与 IgG 的 Fc 片段融合,可延长 IL-10 半衰期,具有低毒性和强大的抗肿瘤活性。它主要通过重新激活和扩增终末耗竭的 CD8 + 肿瘤浸润淋巴细胞(TILs)发挥作用,这些 TILs 通常对免疫检查点抑制剂(ICIs)治疗无反应,而 IL-10/Fc 融合蛋白与 ICIs 或嵌合抗原受体 T 细胞(CAR T)疗法联合使用时,可显著增强抗肿瘤效果,在小鼠的黑色素瘤、卵巢癌和结直肠癌模型中已得到验证。双特异性融合蛋白如 CmAb-(IL-10)₂,将抗 EGFR 抗体(Cetuximab)与 IL-10 二聚体融合,可同时结合 EGFR 阳性肿瘤细胞和 IL-10R,有效延长 IL-10 半衰期,靶向 EGFR 阳性肿瘤,抑制 DC 介导的 CD8+ TILs 凋亡,克服 ICIs 治疗耐药性,与 ICIs 联合使用时展现出协同抗肿瘤效应。BF10 融合蛋白则将 IL-10 与抗 CSF-1R 抗体结合,可靶向 TAMs 富集的肿瘤,同时递送 IL-10 至 TME 并抑制 CSF-1R 信号,消耗 TAMs 并重新编程 CD8+ TILs,在多种癌症模型中展现出显著的抗肿瘤活性,与 ICIs 联合使用可进一步增强疗效。 - 工程化变体
通过生物工程技术改造 IL-10,使其对 IL-10Rβ 的结合亲和力增强。这种变体在低剂量下就具有强大的免疫调节作用,能更有效地招募 IL-10Rβ 形成活性信号复合物,增强 STAT1 和 STAT3 信号通路的激活,进而提高人 CD8+ T 细胞的转录活性。在培养的 CAR T 细胞中,与野生型 IL-10 相比,该变体可使 CAR T 细胞对急性髓系白血病细胞的细胞毒性显著增强。 - 载体递送系统
溶瘤病毒(OVs)如携带 IL-10 基因的溶瘤痘苗病毒(OVV-IL10),利用 OVs 能选择性在肿瘤细胞中复制并裂解肿瘤细胞的特性,同时释放 IL-10,调节机体的先天和适应性免疫反应,刺激抗肿瘤细胞毒性 CD8+ T 细胞,在胰腺癌模型中展现出持续的肿瘤抑制作用,降低疾病复发风险。纳米颗粒和支架等递送系统可将 IL-10 包裹或结合其中,实现 IL-10 的靶向递送和控释,延长其半衰期,增强储存稳定性和抗肿瘤活性,还可实现多种治疗剂的共递送,产生协同治疗效果 。
- 临床试验成果
-
在早期临床试验中,pegilodecakin 单药治疗对部分晚期实体瘤患者有一定抗肿瘤活性,联合抗 PD-1 治疗(如 pembrolizumab 或 nivolumab)在非小细胞肺癌、肾细胞癌等患者中,客观缓解率(ORR)、无进展生存期(PFS)和总生存期(OS)等指标优于单药治疗 。 - 面临的挑战
在后续的 CYPRESS 和 SEQUOIA 临床试验中,pegilodecakin 联合治疗在转移性非小细胞肺癌和转移性胰腺癌患者中未显著改善 ORR、PFS 和 OS,且血液学不良反应发生率较高。这表明 IL-10 介导的肿瘤治疗在不同癌症类型、患者群体和治疗方案组合中存在疗效差异,可能与肿瘤异质性、患者个体差异以及治疗方案的优化程度有关。此外,PEG 化虽有优势,但也可能带来剂量相关的毒性等问题。 - 未来展望
未来 IL-10 介导的肿瘤治疗需要进一步优化递送机制,提高 IL-10 在肿瘤部位的特异性积累和有效浓度,减少对正常组织的副作用。通过深入研究肿瘤生物学和免疫微环境,结合生物标志物进行精准患者分层,筛选出最能从 IL-10 治疗中获益的患者群体。同时,不断探索 IL-10 与其他免疫调节药物、化疗药物或靶向药物的联合治疗方案,以充分发挥 IL-10 的治疗潜力,提高肿瘤治疗的效果和患者生存率。