


一、脂肪组织:从能量储存库到代谢调控中心


脂肪组织曾长期被视为被动的能量储存器官。近年研究表明,它是一个活跃的内分泌与代谢调控中心,在能量稳态、胰岛素敏感性及全身代谢调控中发挥关键作用。根据其形态、功能与分布,脂肪组织主要分为以下三类:



白色脂肪组织(White Adipose Tissue, WAT):广泛分布于皮下和内脏周围,主要功能是以甘油三酯形式储存多余能量。研究中常以iWAT(腹股沟白色脂肪组织)和eWAT(附睾白色脂肪组织)作为模型。
iWAT:属于皮下脂肪,具有更强的脂肪储存缓冲能力及一定的“代谢健康”特性,在特定条件下可发生“米色化”,具备产热潜能;



棕色脂肪组织(Brown Adipose Tissue, BAT):主要分布于肩胛区、颈背部及纵隔周围,细胞内富含线粒体及产热关键蛋白解偶联蛋白1(UCP1),通过非颤抖性产热消耗能量,参与体温维持与能量平衡调节。



米色脂肪组织(Beige/Brite Adipose Tissue):散在于白色脂肪中,并非独立的解剖结构。其在寒冷、运动或激素刺激下可被激活,表达UCP1并获得类似棕色脂肪的产热能力。这一“白色脂肪棕色化”过程被视为治疗肥胖及相关代谢性疾病的潜在靶点。



脂肪组织不仅储存能量,更通过分泌脂肪因子(如瘦素、脂联素)、外泌体等活性物质,与大脑、肝脏、肌肉等器官形成复杂的信号网络,精密调控全身糖脂代谢与免疫稳态。因此,实现对脂肪组织基因功能的精准调控,对于揭示代谢疾病机制及开发新型治疗策略至关重要。然而,传统研究方法(如原代脂肪细胞培养困难、转基因动物构建周期长、成本高)存在诸多局限,促使研究者寻求更高效、灵活的体内基因递送工具。其中,重组腺相关病毒(AAV)载体凭借其高安全性、持久表达和组织特异性转导能力,在脂肪组织研究中展现出独特且强大的优势。


二、AAV载体在脂肪研究中的核心优势
高安全性与持久表达:重组AAV致病性低、免疫反应弱,外源基因可在脂肪细胞中持续表达数月至数年,适合长期代谢观察。
靶向高效且具组织特异性:不同血清型对脂肪组织具有天然趋向性差异。如AAV8和AAV9经局部或全身给药后可高效转导白色与棕色脂肪;工程化血清型Rec2在局部注射中表现更优,且肝脏脱靶显著降低。
精细的表达调控能力:结合组织特异性启动子(如aP2、Adipoq、UCP1等)及miRNA靶序列,可实现基因在特定脂肪细胞类型中的精准表达,并有效抑制非靶组织中的表达。
灵活的功能操控:可承载基因过表达、RNA干扰沉默、CRISPR基因编辑等多种工具,满足从功能验证到疾病建模的多元需求。

三、AAV靶向脂肪组织的策略与方法 为实现脂肪组织的高效、特异性基因递送,研究人员已发展出一套多层次的优化策略,涵盖血清型筛选、启动子选择、以及表达调控元件的设计: 
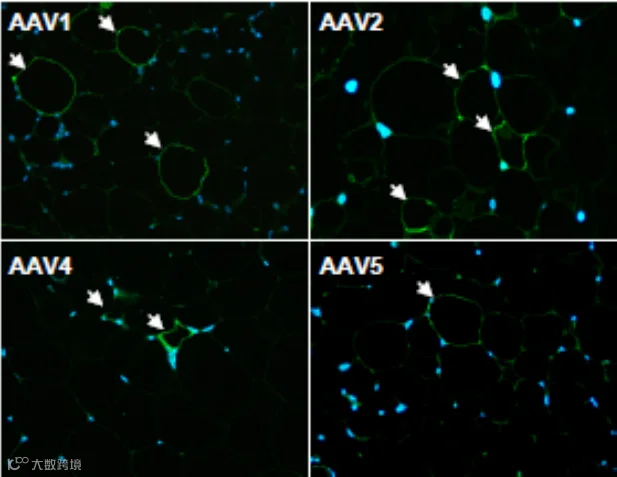
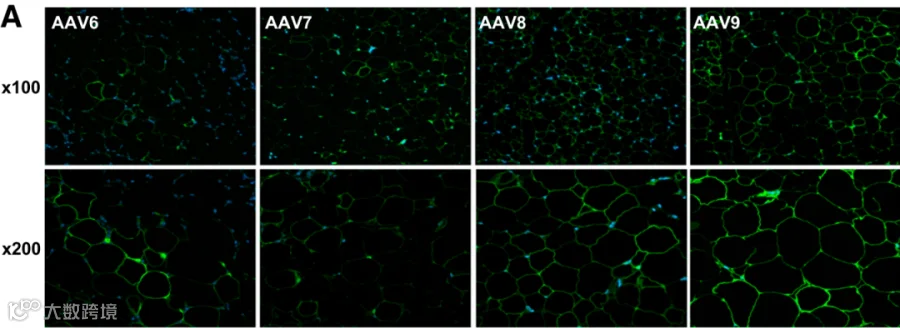
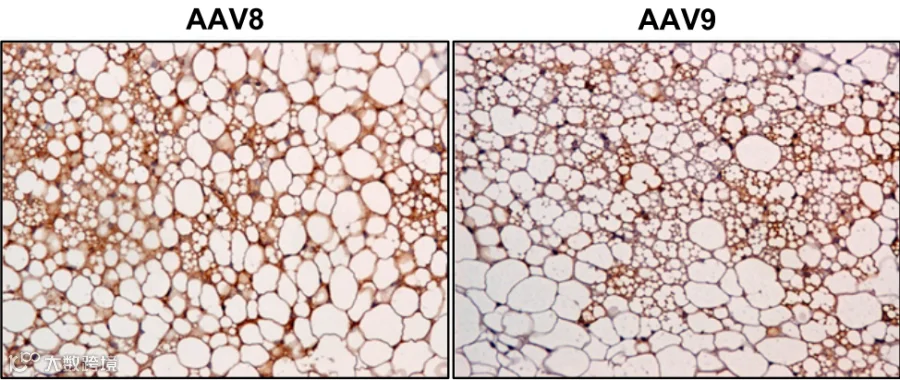
2013年,Jimenez 等人[1]系统评估了多种AAV血清型,发现通过局部注射,AAV8和AAV9在eWAT、iWAT和肩胛间棕色脂肪(iBAT)中的转导效率显著高于AAV1, 2, 4, 5, 6, 7,能够在脂肪细胞中实现广泛且高水平的转基因表达(图1-3)。
图1 eWAT局部注射不同AAV血清型结果
图2 iWAT局部注射AAV8和9, 对iWAT切片进行GFP(棕色)免疫染色
图3 AAV载体经iBAT内 给药后BAT的转导

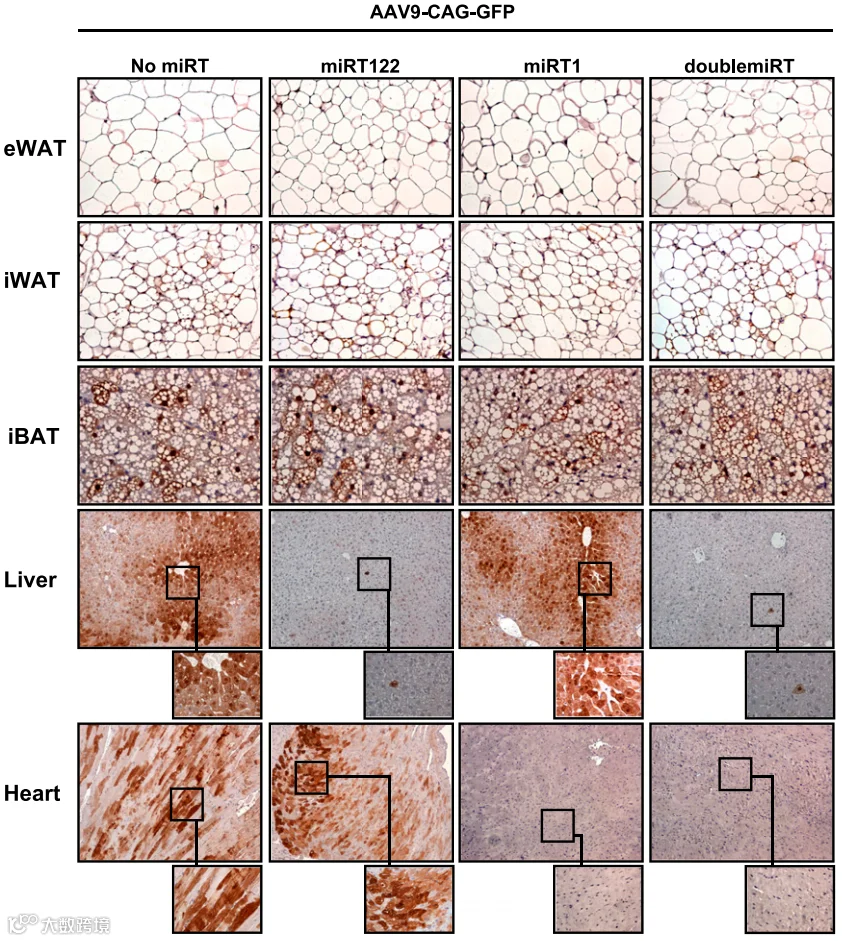 
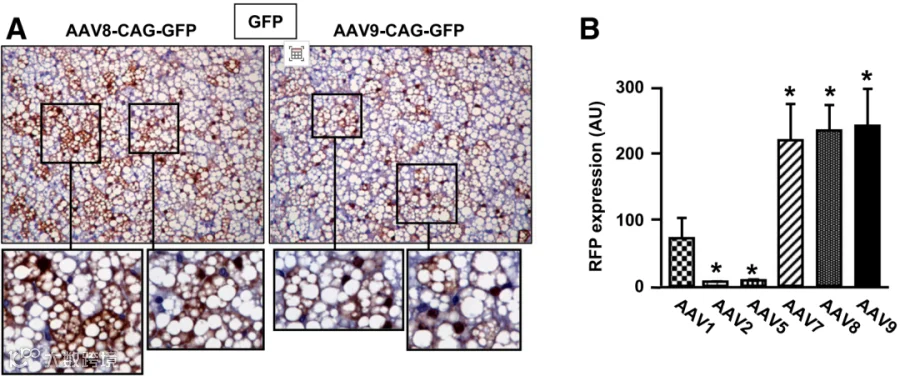
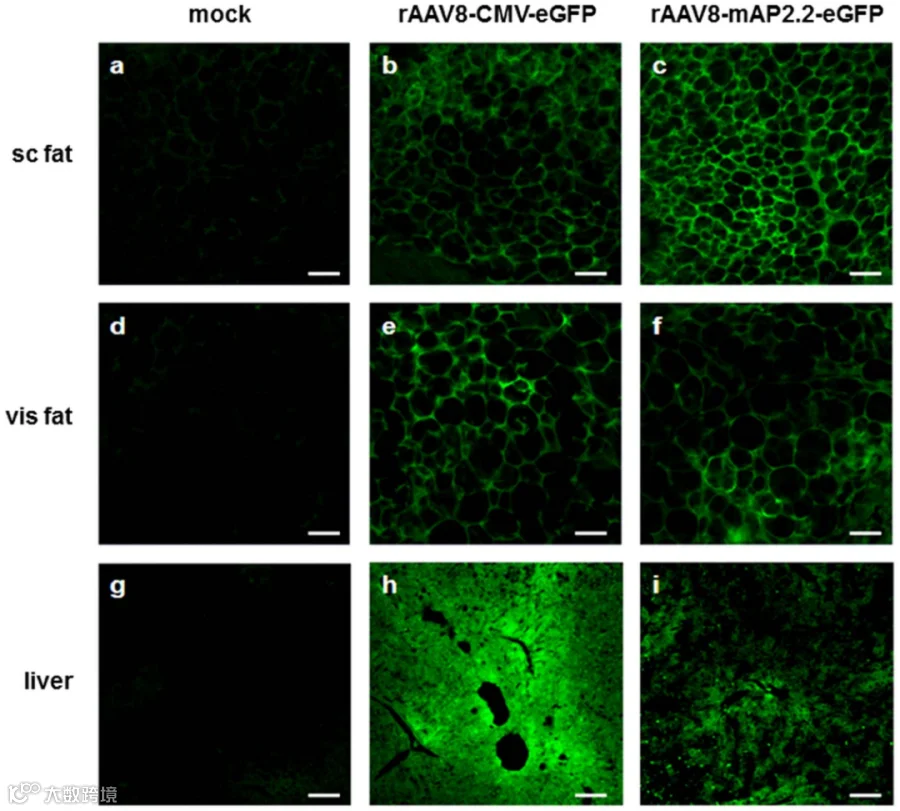 图5 rAAV8-CMV-eGFP和 rAAV8-mAP2.2-eGFP, eGFP表达的组织学评估

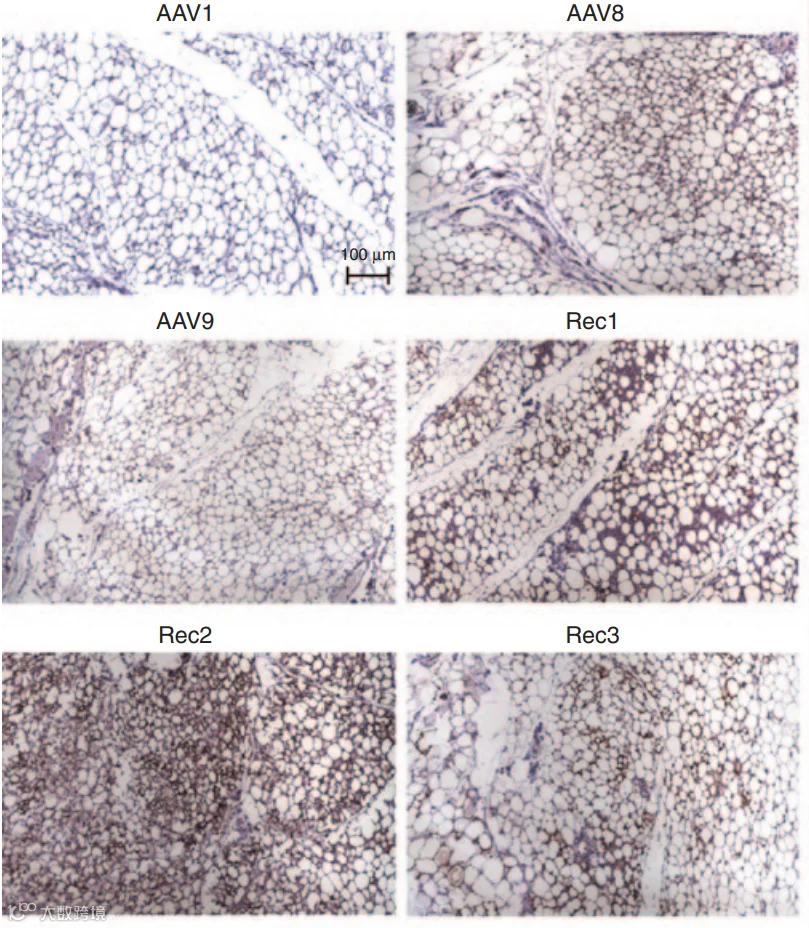
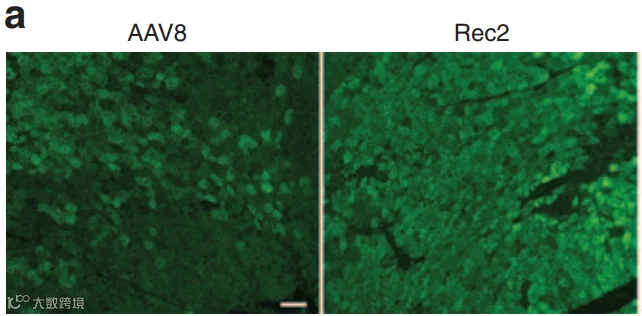
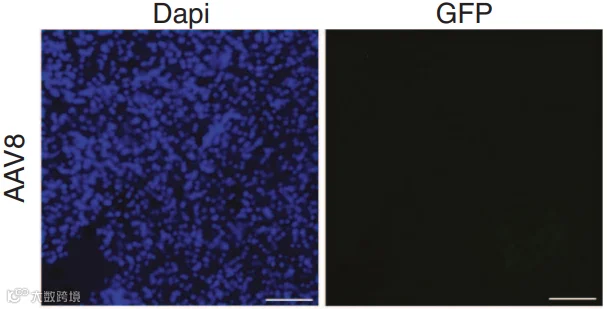
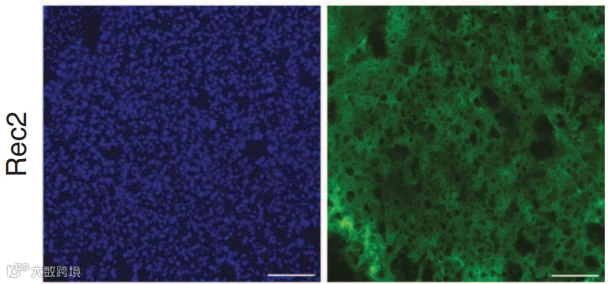
2014年,Liu等人[4]改造的工程化血清型Rec2在脂肪靶向中显示出独特优势。研究发现,Rec2在局部注射至脂肪垫后,尤其在棕色脂肪中的转导效率显著高于AAV1、8、9(图6-7)。(iWAT注射的病毒量为1E+10VG;iBAT为1E+9VG/每侧)
此外,Rec2的给药途径灵活,其组织嗜性受给药方式显著影响[5,6]:
尾静脉注射:Rec2主要转导肝脏,脂肪组织中表达较低;
腹腔注射:Rec2能高效靶向内脏脂肪(如附睾白色脂肪组织eWAT和肠系膜白色脂肪组织mWAT),而皮下脂肪和棕色脂肪中表达极微,且所需剂量比既往全身给药报道低1–2个数量级;
口服给药:一个有趣的发现是,口服Rec2能高效、特异地转导棕色脂肪,为无创研究BAT功能提供了新思路,但该方式在研究中较少作为常规给药途径,更多用于特定机制探索(图8);
局部注射:无论是白色脂肪还是棕色脂肪,Rec2均表现出优异的转导效率。
2025年,该团队[7]进一步对Rec2进行定点突变(F503Y、Y708D和K709I)得到V7血清型,该血清型在全身给药时肝脏嗜性减弱,腹腔注射有利于内脏脂肪的转导,而静脉注射则有利于皮下脂肪,再次表明给药途径显著影响AAV衣壳的生物分布。实验结果显示,皮下注射V7-LEP能高效、特异性地感染皮下脂肪组织,有效恢复循环瘦素水平,并完全纠正ob/ob小鼠的代谢异常,治疗效果可持续至少9个月。 
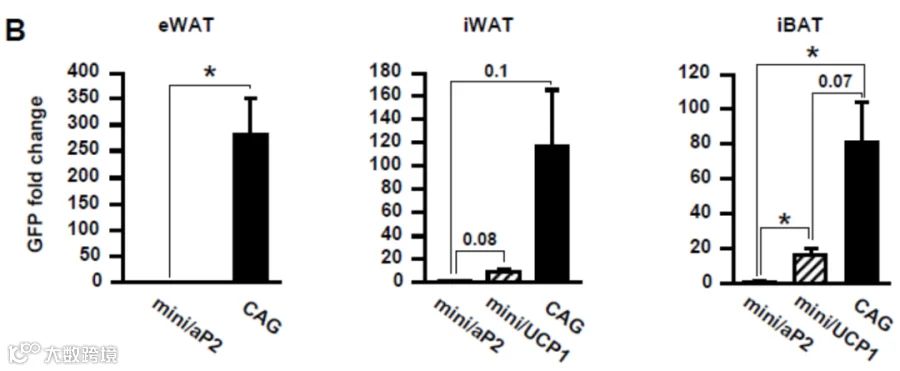
利用脂肪特异性启动子可实现转基因的细胞类型限制性表达: 白色脂肪常用启动子:mini/aP2
Lagarrigue等人[8]通过尾静脉注射AAV8-mini/aP2-cre。病毒基因组检测显示,感染主要局限于BAT、eWAT、皮下白色脂肪组织(s.c. WAT)及肝脏,而在胰腺和骨骼肌中未见明显感染。RT-qPCR分析显示eWAT和s.c. WAT中Cdk4 mRNA显著减少,而肝脏和BAT中无变化。
aP2:Adipocyte Fatty Acid Binding Protein 2, 脂肪细胞脂肪酸结合蛋白2,也称为FABP4。
需注意的是,mini/aP2启动子表达强度很弱,如需使用,建议采用局部注射,高剂量方法提高表达效果。
图9 静脉注射不同AAV载体后, eWAT、iWAT和iBAT中的GFP表达水平 
|
棕色/米色脂肪常用启动子:mini/UCP1
自1998年Cassard-Doulcier[9]验证mini/UCP1对于棕色脂肪组织的特异性表达以来。已有许多研究通过局部注射AAV8/9-mini/UCP1-GFP至iBAT,实现了棕色脂肪细胞的特异性转导,且几乎无心脏表达和极微弱的肝脏表达。如Behrens J等人[10]利用将该启动子、Rec2和miR122靶序列的AAV载体,通过尾静脉注射实现棕色脂肪特异性表达,以及避免肝脏表达的脱靶问题。
广谱脂肪细胞启动子:当研究对象为成熟脂肪组织,或追求更强的表达水平时,小鼠脂联素(mAdiponectin)启动子全长mAP2.2或人源脂联素启动子/增强子hAdp[14]或是更佳选择。例如,Uhrig-Schmidt等[2]在体外验证实验中发现mAP2.2启动子在未分化的前脂肪细胞中表达eGFP水平显著较低,分化成熟的脂肪细胞中,eGFP表达与内源性脂联素表达同步显著升高;Sancar等人[11]在研究FGF1调控脂解的机制时,采用AAV8搭载hAdp启动子,成功在成熟脂肪细胞中实现了PDE4D3的特异性过表达,未验证在前体脂肪细胞的表达效率。



四、AAV在脂肪研究中的典型应用实例









五、总结 |
应用前景
基因治疗平台:利用脂肪组织易获取、具分泌能力的特点,可将其改造为“体内生物工厂”,长效生产治疗性蛋白,用于糖尿病、血脂异常等全身性疾病。
直接治疗策略:通过递送UCP1、FGF21等基因诱导白色脂肪棕色化、增强能量消耗,为肥胖及相关代谢病提供新途径。
未来展望
实验方案建议
01
白色脂肪组织
首选血清型:AAV8 或 Rec2。
推荐启动子:mini/aP2:适用于白色脂肪细胞特异性表达。
给药途径与剂量参考:
局部注射(iWAT或eWAT):1×10¹⁰ – 2×10¹¹ vg。
尾静脉注射(全身靶向):1×10¹² – 5×10¹² vg。
02
棕色脂肪组织
首选血清型: Rec2或 AAV8/AAV9。
推荐启动子:mini/UCP1:实现棕色脂肪高特异性表达。
给药途径与剂量参考:
局部注射(肩胛间BAT):1×10⁹ – 2×10¹⁰ vg。
03
通用优化策略
降低脱靶:在载体3‘UTR中插入肝脏miR-122靶序列(如4×或8×重复),可显著抑制肝脏表达。
剂量调整提示:使用组织特异性启动子(如mini/aP2、mini/UCP1)时,其驱动效率低于广谱启动子(如CAG/CMV),通常需要提高5–20倍剂量以达到同等表达水平。
注:注射剂量为文献中所用剂量,考虑实际条件区别,建议客户做预实验探究最适剂量。



Taitool现货列表

货号
|
产品名称
|
S1096-9
|
AAV2/9-hAdp-EGFP-P2A-iCre-miR122Det-WPRE-pA
|
S0751-9
|
AAV2/9-hAdp-EGFP-WPRE-pA
|
S1142-9
|
AAV2/9-hAdp-MasterRNAi155(NC)-miR122Det-mCherry-WPRE-pA
|
S2014-8
|
AAV2/8-mini/aP2-Cre-miR122Det-WPRE-pA
|
S2015-8
|
AAV2/8-mini/UCP1-Cre-miR122Det-WPRE-pA
|
S2016-8
|
AAV2/8-mini/UCP1-EGFP-miR122Det-WPRE-pA
|
S2017-8
|
AAV2/8-mAP2.2-EGFP-miR122Det-WPRE-pA
|
[1]Jimenez V, Bosch F, et al. In vivo adeno-associated viral vector-mediated genetic engineering of white and brown adipose tissue in adult mice. Diabetes. 2013 Dec;62(12):4012-22.
[2]Uhrig-Schmidt S, Kreuz S, et al. Gene delivery to adipose tissue using transcriptionally targeted rAAV8 vectors. PLoS One. 2014 Dec 31;9(12):e116288.
[3]Mizukami H, Ozawa K, et al. Adipose tissue as a novel target for in vivo gene transfer by adeno-associated viral vectors. Hum. Gene Ther. 2006;17:921–928.
[4]Liu X, Cao L, et al. Adipose tissue insulin receptor knockdown via a new primate-derived hybrid recombinant AAV serotype. Mol Ther Methods Clin Dev. 2014 Feb 5;1:8.
[5]Huang W, Cao L, et al. Genetic Manipulation of Brown Fat Via Oral Administration of an Engineered Recombinant Adeno-associated Viral Serotype Vector. Mol Ther. 2016 Jun;24(6):1062-1069.
[6]Huang W, Cao L, et al. Targeting Visceral Fat by Intraperitoneal Delivery of Novel AAV Serotype Vector Restricting Off-Target Transduction in Liver. Mol Ther Methods Clin Dev. 2017 Jun 19;6:68-78.
[7]Cao L, Huang W, Xiao M, Zou X. Subcutaneous administration of adipose-tropic gene therapy for congenital leptin deficiency. Res Sq [Preprint]. 2025 Jul 17:rs.3.rs-7030870..
[8]Lagarrigue S, Fajas L, et al. CDK4 is an essential insulin effector in adipocytes. J Clin Invest. 2016 Jan;126(1):335-48..
[9]Cassard-Doulcier AM, Ricquier D, et al. A 211-bp enhancer of the rat uncoupling protein-1 (UCP-1) gene controls specific and regulated expression in brown adipose tissue. Biochem J. 1998 Jul 15;333:243-6.
[10]Behrens J, Scheja L, et al. An efficient AAV vector system of Rec2 serotype for intravenous injection to study metabolism in brown adipocytes in vivo. Mol Metab. 2024 Oct;88:101999.
[11]Sancar G, Evans RM, et al. FGF1 and insulin control lipolysis by convergent pathways. Cell Metab. 2022 Jan 4;34(1):171-183.e6.
[12]Tiwari M, Mcilroy GD, et al. Preclinical evaluation of tissue-selective gene therapies for congenital generalised lipodystrophy. Gene Ther. 2024 Sep;31(9-10):445-454.
[13]McCann MA, Liew CW, et al. Adipose expression of CREB3L3 modulates body weight during obesity. Sci Rep. 2021 Sep 29;11(1):19400.
[14]O'Neill SM, Reilly MP, et al. Targeting adipose tissue via systemic gene therapy. Gene Ther. 2014 Jul;21(7):653-61. doi: 10.1038/gt.2014.38. Epub 2014 May 15. PMID: 24830434; PMCID: PMC4342115.
END


扫码了解
更多信息

点击蓝字关注我们吧!!