
From Engram to Neural Ensemble, Memorandum of Activity Dependent Tools
随着科技及产业巨头纷纷锚向“人工智能(Artificial Intelligence)”,一时间AI大热。然而任何技术(算法构造)的革新都无法湮没一个近乎事实的断言:伟大的人工智能必需建立在神经科学(Neuroscience)的基础上,而回归神经科学的核心无疑是破译大脑的工作机制(Hassabis et al. 2017)。

当前细胞及环路水平的研究认为,大脑的功能编码很可能是存在于特定的神经类群激活/抑制的时间空间图式(Neural activity patterns)之中。解析功能编码方面活跃着两大重要的指导理论(Tonegawa et al. 2015):
20世纪初叶,德国进化生物学家西蒙(Richard Semon)开创性地提出了印迹理论(Engram Theory)阐述记忆编码:记忆形成和存储于机体被相应刺激所同步激活的、发生了修饰变化的、分离却联合统一的激活复合体(印迹)之中,而随后学习刺激经历的部分线索/境况即可将之再度激活引起记忆的召回(Semon. 1904, 1909)。

20世纪中叶,加拿大杰出的神经心理学家赫布(Donald O. Hebb)在其经典著作《行为的组织》(The organization of behavior)中提出了神经集群(Cell Assembly/NeuralEnsemble)的概念用以描述突触可塑性理论(Synaptic Plasticity Theory),这一被环境信号特异集结激活的,特定模式的神经类群构成了编码处理学习偶联信息的单元模块(Hebb. 1949),而神经集群间突触连接的强化被认为是形成印迹所必需的(Josselyn et al. 2015)。
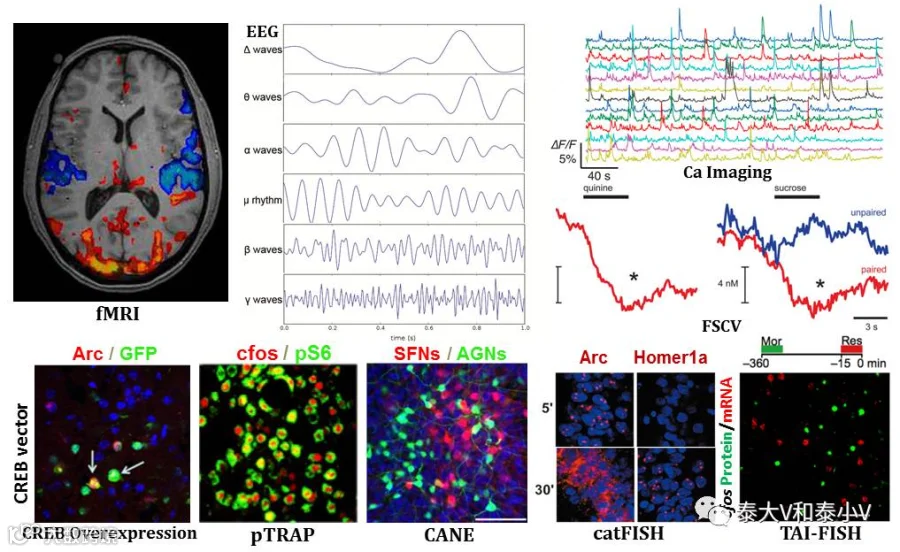
要想破译及模拟大脑的功能编码势必要先实现大脑活动的检测。检测脑区及神经元活化的方法众多:有基于代谢的2-脱氧葡萄糖(2-DG)摄取的放射自显影(Sokoloff et al. 1977)、近红外光谱技术(NIRS)(Jobsis et al. 1977)、血氧水平依赖的功能核磁共振(BOLD-fMRI)(Ogawa et al. 1990)及正电子发射断层成像(PET/SPECT)(Kato et al. 1990; Bryant etal. 1998);基于电/磁信号的脑电图(EEG)(Berger H, 1929)/脑磁图(MEG)(Cohen D. 1968)记录(da Silva F L, 2013)以及电生理记录(Patch clamp/In vivoRecording)(Neher & Sakmann,1976;Hubel, 1960;McNaughton et al. 1983);基于钙信号的光纤记录(Fiber Photometry)(Adelsberger et al. 2005;Cui et al. 2013)、双光子钙成像(Two-photon Ca imaging)(Trachtenberg et al. 2002;Grutzendler et al. 2002)以及显微内窥镜(Grinlen/Microendoscope)(Hamel et al. 2015);检测神经化学变化的微透析/快速扫描循环伏安法(Microdialysis/FSCV)(Ungerstedt et al. 1974;Robinson et al. 2003);还有检测即早基因(IEG)cfos(Morgan et al. 1987),Arc/Arg3.1(Link et al. 1995; Lyford etal. 1995),zif268/egr1(Saffen et al. 1988),homer1a(Brakeman et al. 1997; Kato etal. 1997)等的表达以及转录因子CREB(环腺苷酸反应元件结合蛋白)(Ginty et al. 1993; Bito etal. 1996)、核糖体蛋白S6的磷酸化(Knight et al. 2012)等。
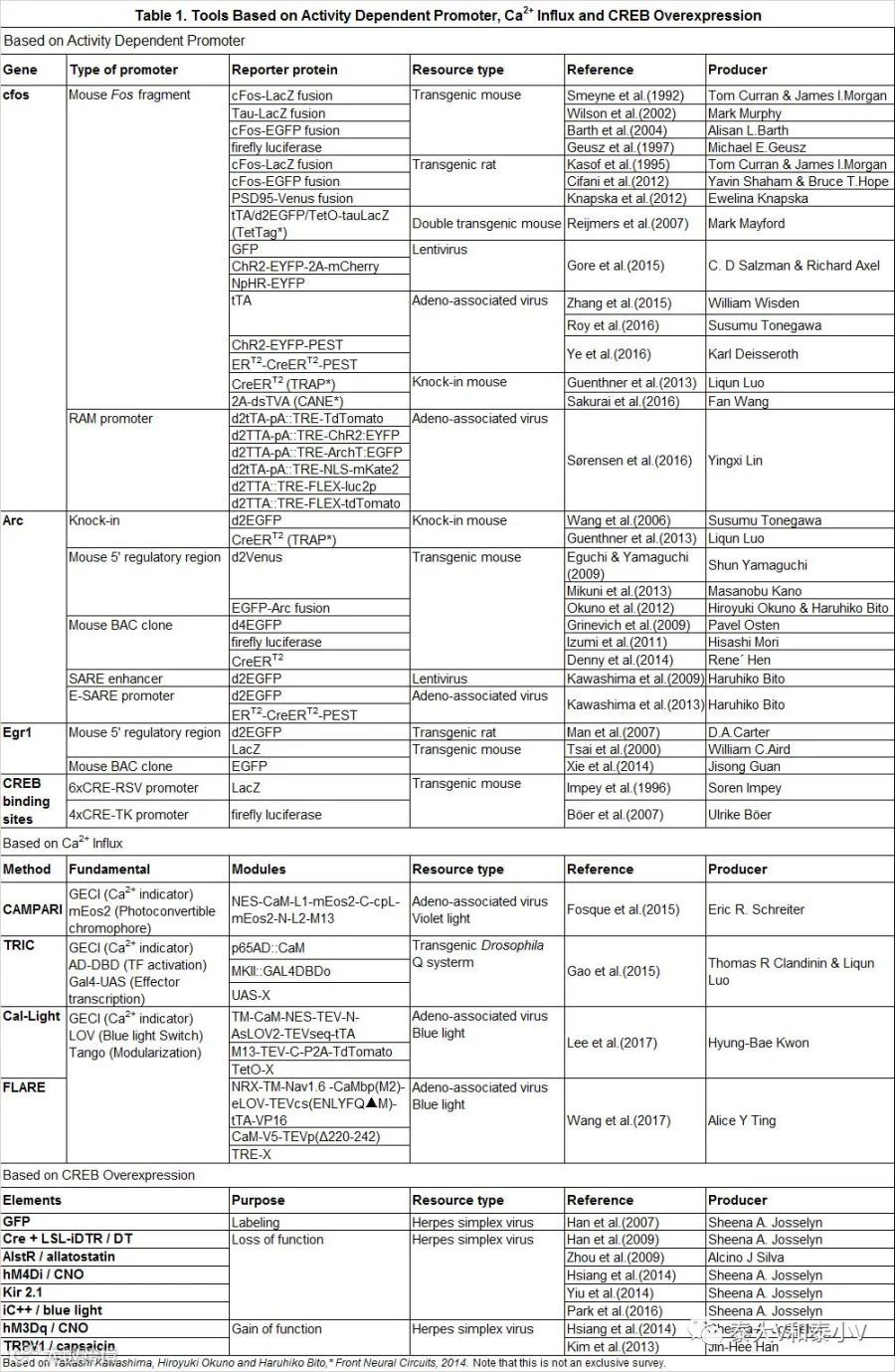
活性检测(Activity visualization)提供了功能解析最基本的相关性证据,然而更深层的功能编码(Encoding)的论断还需要相应的功能获得(Sufficiency)和功能缺失(Necessity)的验证支撑。从经典的电刺激、损毁、药理兴奋/抑制(Agonist/Antagonist)到本世纪大放异彩的光遗传(Optogenetics)/药理遗传(DREADDs)、细胞凋亡/突触抑制(taCasp3/TetTox)、过表达、敲减及沉默(RNAi/CRISPRi)等诸多干预的方法为解析脑区、细胞及环路等的功能编码开拓了道路,但大脑远比我们所见过的任何机器都要精密复杂,脑区及细胞存在很大的异质性(Heterogeneity),常常无法使用特定的标记(核团亚区、投射及细胞类型)分选出功能类群,于是乎直接捕获,尔后操纵/干预,事件/刺激所激活的神经类群,亦即上述的印迹/神经集群,在神经科学界可谓一直令人心驰神往。自上世纪90年代以来,一系列活动依赖的遗传改造实验大鼠、小鼠及病毒工具被相继开发应用(见表1)(Kawashima et al. 2014)。
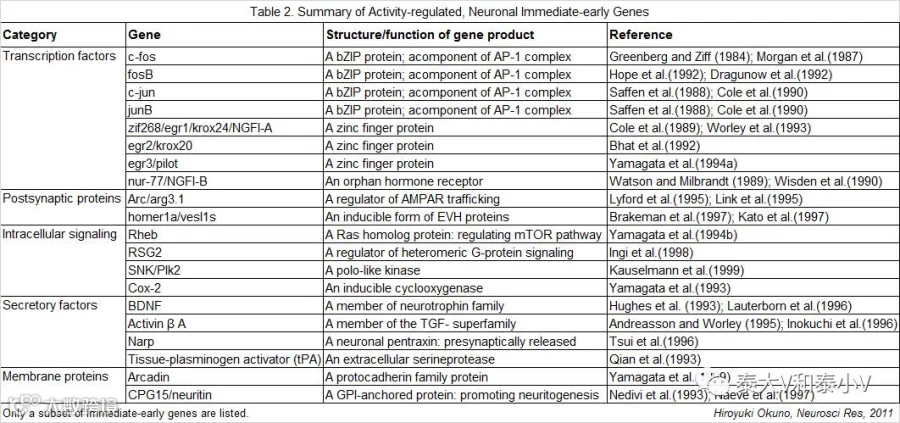
细胞接受外界刺激激活通常会引发一系列的反应,从即时的离子通道开放、Ca2+内流、神经元动作电位、即早基因表达到G蛋白偶联/酶联受体激活级联的长程反应及基因分子水平的变化等生理/生化事件(Morgan & Curran. 1989)。其中即早基因作为细胞活化的分子标记被青睐已久。这一起源于病毒学的概念(Watson & Clements. 1980)自提出以来很快被引入细胞研究(Kelly et al. 1983;Greenberg & Ziff. 1984)。IEG在受到外界刺激后迅速启动转录,不依赖于新的蛋白合成。自上世纪80年代末被引入神经科学研究,分离解析新的神经IEG用于神经可塑性及认知功能解析(尤其是探索学习记忆的“主控基因”)以及应用IEG的表达(mRNA/Protein)表征大脑神经活性的研究层出不穷,大量的即早基因被发现,其表达产物涵盖了转录因子、突触后蛋白、信号分子、分泌因子及膜蛋白(见表2)(Ouno. 2011)。
c-myc和c-fos是最早被鉴明的即早基因(Sheng & Greenberg. 1990)。cfos表达的分子机制如下图所示:
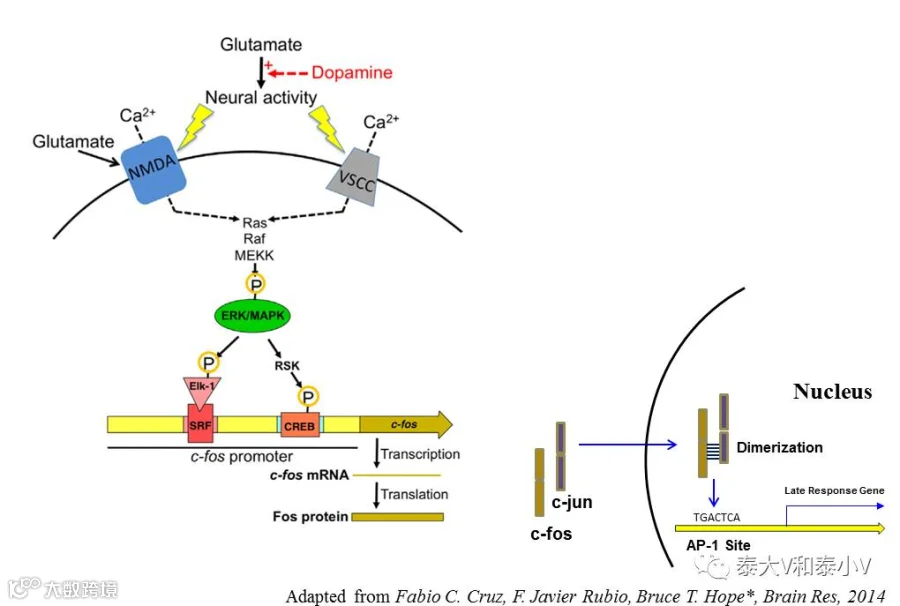
除了cAMP/PKA通路的激活,神经元活化引发了NMDAR及VSCC介导的Ca2+内流,当胞内Ca2+达到一定的水平会导致磷酸化及通过ras-raf-MEKK级联信号通路激活ERK/MAPK,最终致使转录因子Elk-1/SRF 及CREB磷酸化激活,从而结合在cfos基因的5’非编码调控区(SRE/CRE/CaRE)启动cfos表达(Cruz et al. 2015)。生成的转录因子Fos与同家族的转录因子Jun通过亮氨酸拉链形成异二聚体,随后识别并结合在含有AP-1位点的基因上调控晚期基因的转录(Curran & Morgan.1995)。
另一被广泛研究的即早基因Arc与突触活动高度相关,其产物是一个富集于突触后致密区(PSD)的突触蛋白,能够通过与细胞骨架蛋白相互作用而强化膜受体的内吞作用(AMPAR trafficking),被认为参与了突触标度(Synaptic scaling)这一自身稳态的突触可塑性(Okuno. 2011)。调节Arc表达的一系列元件被称为突触活性响应元件(SARE),包含了CREB, MEF2及SRF三个转录因子的结合位点。Arc表达可能的分子机制如下图所示:
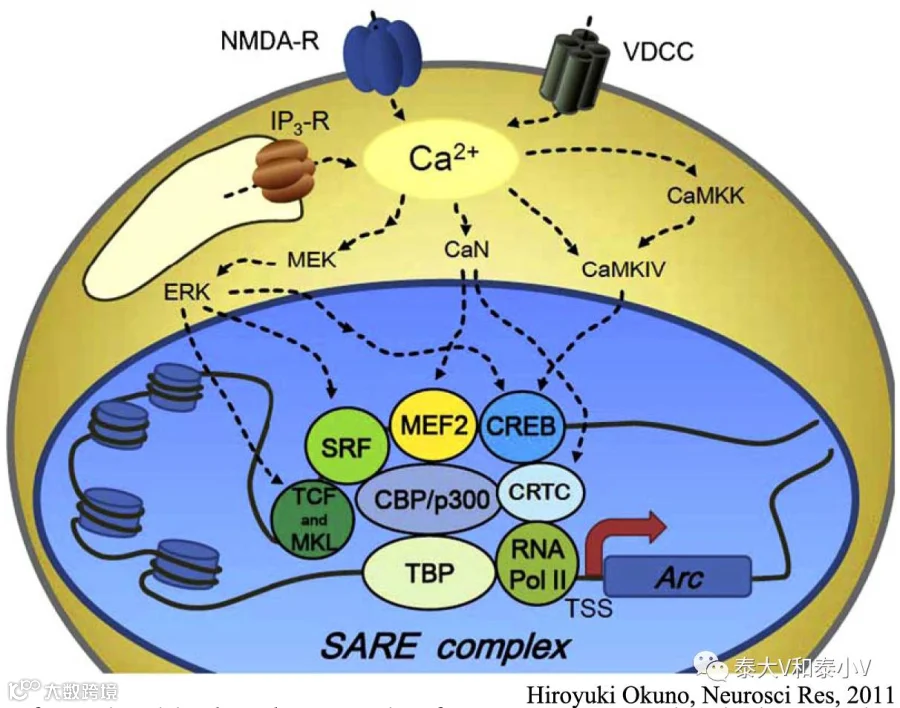
环腺苷酸响应元件(CRE)几乎存在于所有IEG的启动子区域,而CREB被认为是调控IEG表达的一个核心因子。Ca2+内流、钙依赖激酶介导的级联反应及相关转录因子磷酸化活化是IEG表达的基本要素(Kawashima et al. 2014)。但需要注意的是,不同IEG在不同的脑区及细胞类型中表达有所差异,可能是转录调控及钙调动力学上面存在的差异所致(Kawashima et al. 2014),此外有的IEG在某些脑区的表达背景很高,需要根据具体的实验需求进行探索验证。
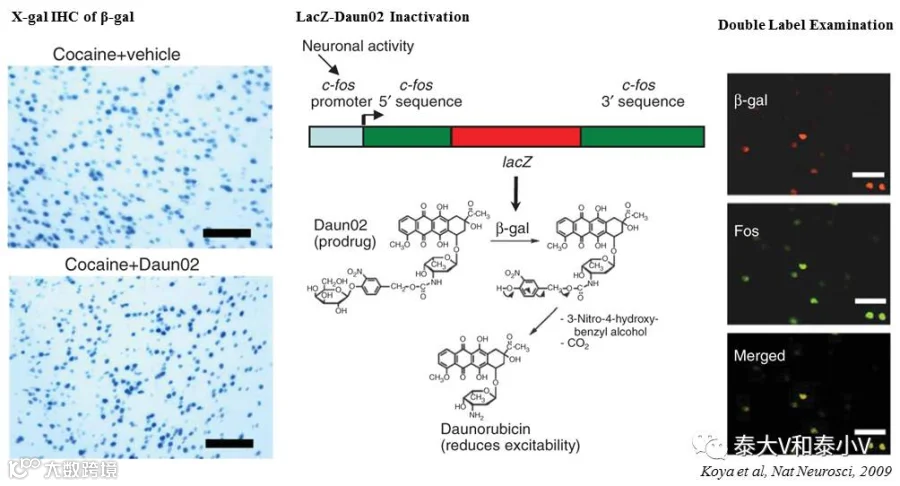
最早开发的活动依赖工具利用IEG直接驱动报告基因如β-半乳糖苷酶(β-gal)基因LacZ(Smeyne et al. 1992,Kasof et al. 1995, Impey et al. 1996, Tsai etal. 2000, Wilson et al. 2002)、荧光素酶(Luciferase)基因(Geusz et al. 1997, Böer etal. 2007, Izumi et al. 2011)及荧光蛋白(GFP/Venus/d2EGFP/d2Venus)基因(Barth etal. 2004, Wang et al. 2006, Man et al. 2007, Grinevich et al. 2009,Eguchi & Yamaguchi. 2009, Cifani etal. 2012, Knapska et al. 2012, Okuno et al. 2012, Mikuni et al. 2013, Xieet al. 2014)的表达来实现活化神经类群的标记。荧光标记的实验动物一方面可以直接用于绘制全脑活性图谱、大批量对比筛选响应刺激的相关脑区及类群(Man et al. 2007, Kim et al.2015, 2016),另一方面可用于脑片电生理研究激活类群的生理特性(Barth et al. 2004, Grinevichet al. 2009)及在体成像研究动态编码(Wang et al. 2006, Xie et al. 2014);荧光素酶报告具有比荧光蛋白报告更高的灵敏度及信噪比,荧光素酶催化底物荧光素(Luciferin)产生的生物发光信号(Bioluminescence)可直接用于非侵入深脑区在体成像(Geusz et al. 1997, Izumi etal. 2011),但该信号的分辨率有限,且时程受底物影响;而LacZ这一经典的报告基因的表达产物也需要借助酶反应(催化底物X-gal显色)才能进一步实现表征(Smeyne et al. 1992, Kasofet al. 1995),但值得一提的是,LacZ也是最早被用于干预的工具之一(稍晚于CREB功能失活工具,见后文)。NIDA的Bruce T. Hope教授利用β-gal能够催化前体药物Daun02转化成具有抑制神经元兴奋作用的柔毛霉素(Daunorubicin)的特性,将Daun02通过套管给予刺激后的cfos-LacZ大鼠的感兴趣脑区,从而实现刺激相关类群的失活(Koya et al.2009,2016)。
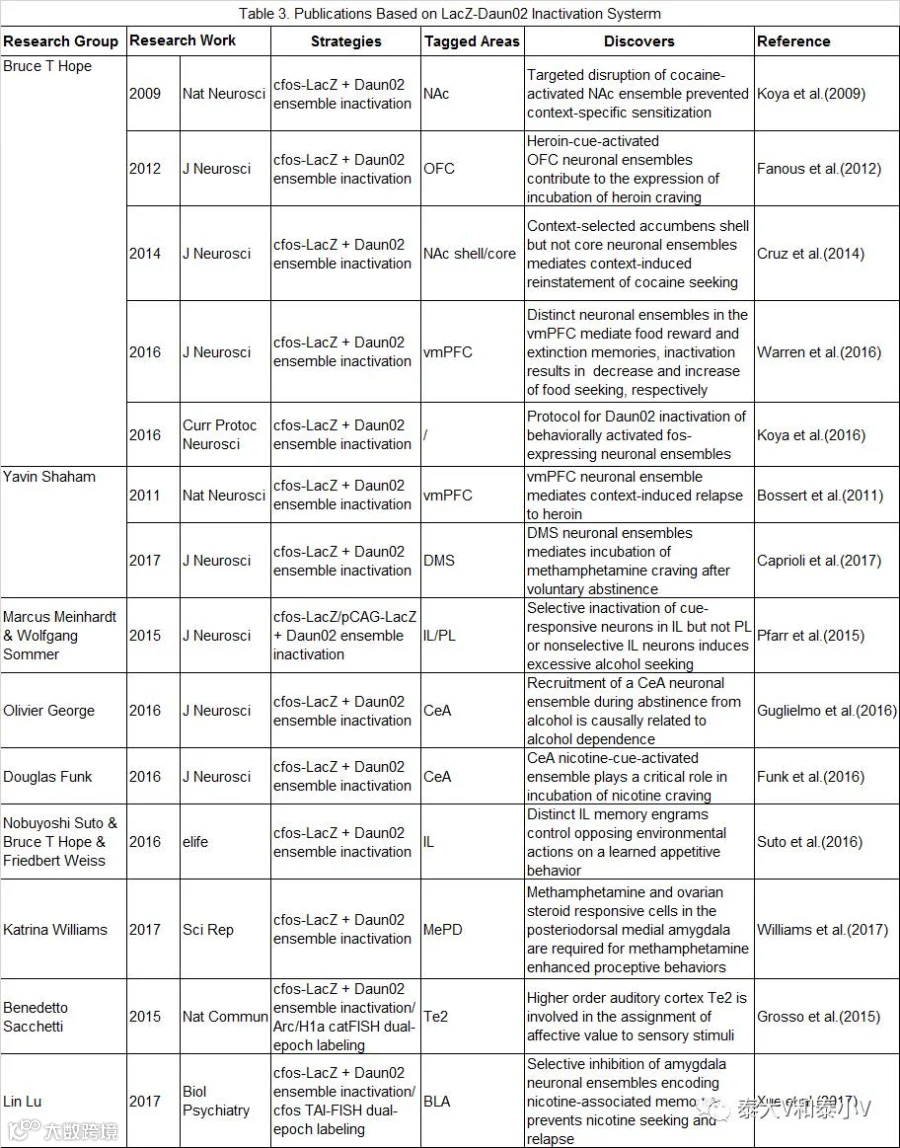
这一方案在研究成瘾药物(Cocaine/heroine/methamphetamine /nicotine/alcohol)偶联环境引起的行为敏化(Sensitization)、渴求(Craving/seeking)、复吸(Relapse/reinstatement)等的机制脑区(NAc/DMS/PFC/OFC/BLA/CeA/MePD/Te2)神经集群的因果作用(Causal role)解析方面有一系列的工作报道(见表3)。
需要注意的是,绝大多数活动依赖的报告动物仅提供了工具特异性的样图,缺乏足够的验证工作,其表达背景及表达模式的活动依赖特性存疑,LacZ动物即因为存在效应蛋白(Effector)相对稳定难降解致使背景偏高的问题而遭受质疑(Kawashima et al. 2014)。
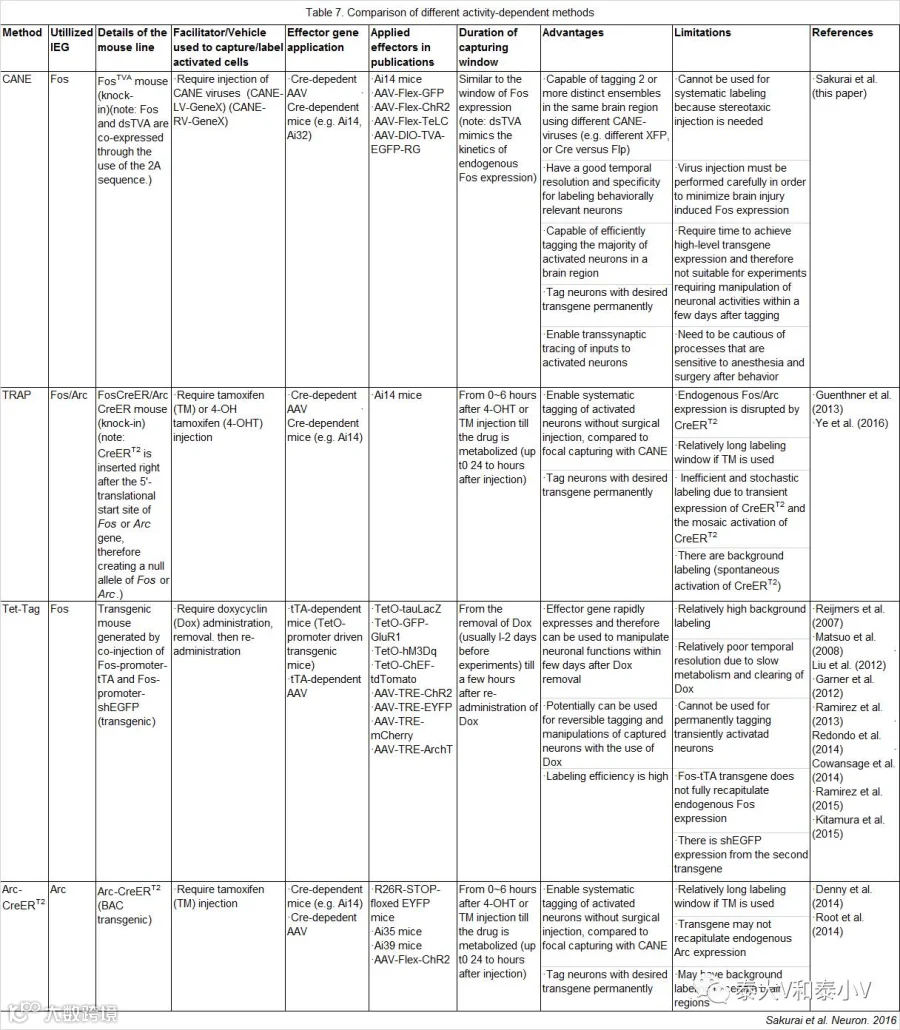
后续的工具则致力于通过引入条件性基因表达控制元件等来改善工具的精度及拓展性,提升活动依赖工具的特异性(Specificity, 捕获与内源激活模式一致的保真信号)、外显率(Penetrance, 捕获尽可能多的保真信号凸显工具效能)及信噪比(S/N ratio, 提升工具灵敏度及降低背景泄漏(Leakage)的综合度量)。目前被开发的有基于Tet-off系统的四环素标签(TetTag)工具;基于CreER诱导重组系统的活性类群靶向重组(TRAP)工具;基于可降解受体dsTVA的活化神经集群捕获(CANE)工具以及基于光控Ca2+依赖的新工具Ca-Light和FLARE,此外还有两个人工改造的活性启动子E-SARE和RAM。在分章简述以上活动依赖工具工作原理及其研究概要之前,我们先对基于CREB过表达工具的募集编码研究作一个简要的综述。
CREB Overexpression
师从加州大学洛杉矶分校(UCLA)Alcino J. Silva教授,多伦多大学病童医院(SickKids)的Sheena A. Josselyn是记忆配置(Memory allocation)研究领域的代表性人物。她和Alcino J. Silva所领导的研究组的一系列研究表明,转录因子CREB水平更高的外侧杏仁核(LA)神经元在恐惧记忆形成过程中会被偏好性激活获得竞争优势,从而被选择性地募集编码记忆印迹(见表4)。
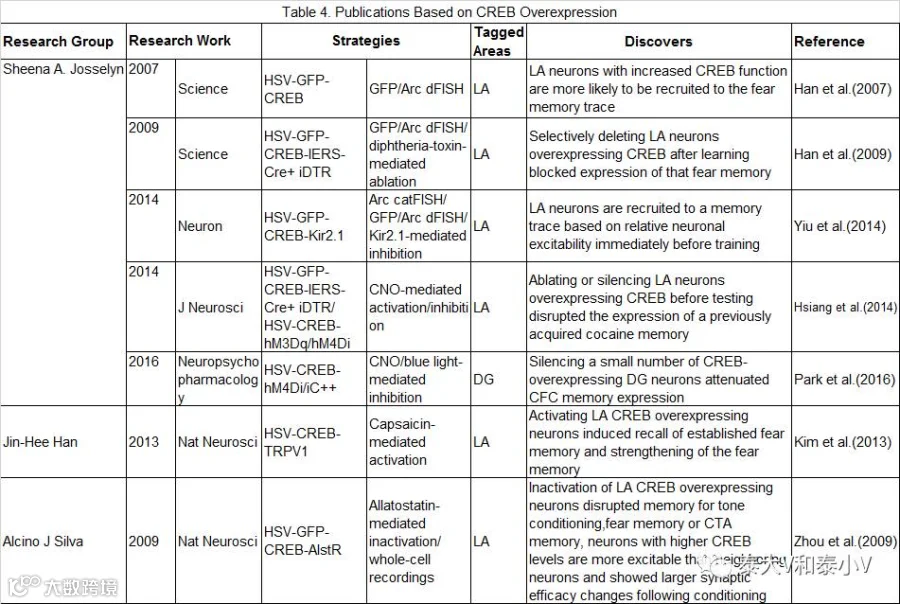
前人的研究表明LA可塑性(Plasticity)对于声音偶联条件性恐惧(Auditory conditioned fear,AFC)记忆是必需的,如同海马(Hippocampus)对于环境偶联条件性恐惧(Context conditioned fear, CFC)记忆的形成。虽然约有70%的LA神经元会接受声音偶联条件性恐惧必要的感觉输入,但只有1/4的LA神经元会发生可塑性变化(Rumpel et al. 2005),被募集编码一次特定恐惧记忆的印迹。早在2007年Sheena A. Josselyn实验室即在《科学》杂志发文证明CREB在恐惧记忆印迹形成的神经元竞争与选择中存在重要作用(Han et al. 2007):他们在CREB缺陷型及野生型(WT)小鼠的LA通过HSV(复制缺陷型单纯疱疹病毒)载体表达融合了GFP的野生型、显性突变型(Dominant-negative)CREB或是空载体对照(感染约20%的LA神经元),并在24h后进行AFC, 结果表明HSV-GFP-CREB成功地恢复(Rescue)了CREB缺陷型小鼠的恐惧记忆,而HSV-GFP-CREB过表达也可增强WT小鼠弱刺激学习(0.4mA电击)引起的恐惧记忆。此外神经元活化分子标签Arc及GFP双重荧光原位杂交(dFISH)实验表明恐惧记忆测试显著地偏好激活GFP-CREB+的神经元,而表达显性突变型CREB的WT小鼠则并没有表现出恐惧记忆的缺陷,但AFC募集编码则会转向GFP—的神经元,结果显示依旧有25%的LA神经元被恐惧记忆测试所激活(Han et al. 2007)。
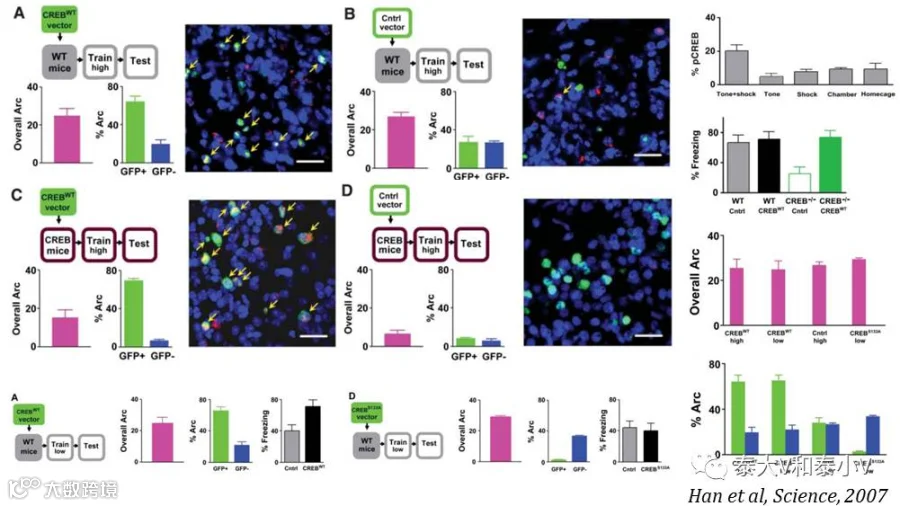
随后发表于2009年的两项工作则首次为记忆印迹编码记忆的必要性提供了直接证据:Sheena A. Josselyn实验室利用可诱导白喉毒素(Diphtheria-toxin, DT)毒杀细胞的策略(HSV-GFP-CREB-IRES-Cre / HSV-GFP-IRES-cre注入iDTR小鼠的LA,AFC之后给予DT)在AFC后将LA 过表达CREB的神经元选择性毒杀,成功地擦除了AFC恐惧记忆。有意思的是这一恐惧记忆虽然被持久擦除但可以被再次习得(亦即募集新的LA神经元编码记忆印迹)(Han et al. 2009)。
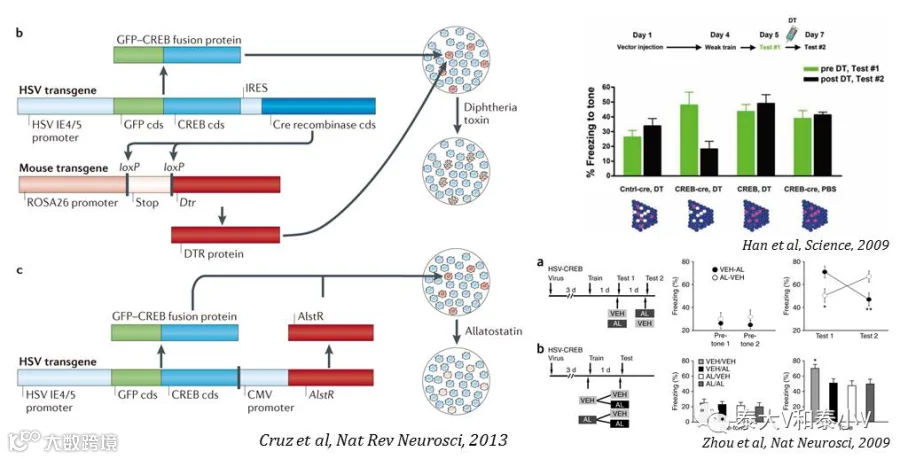
而Alcino J Silva实验室则采用果蝇咽侧体抑制素(allatostatin, AL)G蛋白偶联受体(AlstR)- 配体系统可逆性失活神经元的方案(将HSV-GFP-CREB/LacZ-CMV-AlstR注入小鼠LA,然后进行AFC或是条件性味觉厌恶学习(CTA),AL通过套管局部给到LA)将LA 过表达CREB的神经元选择性失活,可逆且急性地抑制了恐惧记忆及厌恶反应。有趣的是LA表达CREB-AlstR的小鼠不论是在条件性学习(Conditioning)之前半小时还是之后半小时给予AL都会破坏恐惧记忆。此外他们还通过电生理记录证实条件性学习之后过表达CREB的神经元(GFP+)与周围神经元(GFP-)相比兴奋性更高,突触效能(Synaptic efficacy)的改变也更大。他们推测LA里面兴奋性更高的神经集群在学习的过程中可能更容易被激活从而被募集参与记忆编码,另随着CREB激活,CREB抑制因子(Repressor)也可能会被转录从而调整记忆分配尤其是不同记忆的分离(Zhou et al. 2009)。
此后他们又进一步开发了异位大鼠草酸受体TRPV1-辣椒素(Capsaicin)系统,应用这一系统证明激活AFC后LA过表达CREB的神经集群可直接引发恐惧记忆的召回(Recall)及后续的记忆巩固,由于记忆巩固涉及到新的蛋白合成,人工激活同样会致使记忆易变(Lability),他们发现此时给予蛋白合成抑制剂茴香霉素(Anisomycin)会导致相应的恐惧记忆受损(Kim et al. 2013)。随后他们还构建了CREB-Kir2.1内向整流K+通道抑制系统(Yiu et al. 2014)、CREB-DREDDs药理遗传兴奋/抑制系统(Hsiang et al. 2014)及CREB-iC++光遗传抑制系统(Park et al. 2016)进行了一系列解析记忆配置的研究,有意思的是其中一项使用CREB-hM3Dq人工激活可卡因记忆印迹的工作表明人工激活反而削弱了表征可卡因线索记忆的条件性位置偏好(CPP),而作为对照的泛激活组(GFP-HM3Dq)的CPP却不会受影响,与之前人工激活引发恐惧记忆的强化(Kim et al. 2013)不同,这一称之为编码后任意激活(Indiscriminateartificial activation)反而产生了消退(Extinction)的效果(Hsiang et al. 2014);另一项工作则证实海马的齿状回(DG)亚区同样存在募集编码记忆印迹的现象(Park et al. 2016)。
由于LA的募集编码与神经元的相对活性有关(Yiu et al. 2014),后续与LA记忆配置有关的研究则转向了直接使用药理遗传或是光遗传激活提供兴奋类群以纳入记忆印迹(Rashid et al. 2016)。
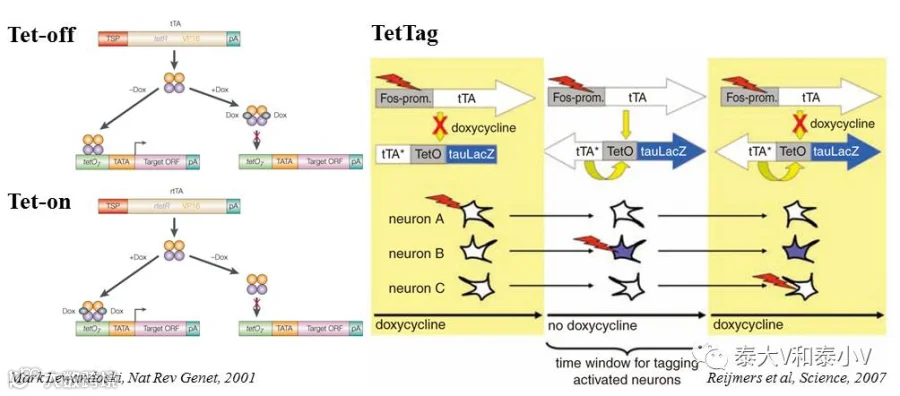
TetTag
四环素标签(TetTag)工具是Scripps研究所的 MarkMayford教授所开发的基于Tet-off调控表达的双转基因活动依赖小鼠。包含了即早基因cfos启动四环素反式激活因子(tTA)及核定位半衰期为两小时的EGFP(shEGFP)表达的活动依赖转基因株系,以及tTA调控(tetO)tau-LacZ表达和非Dox调控组成活性型tTA*表达的反馈环持久标记转基因株系两个部分(Reijmers et al. 2007)。TetTag及后续分离出来的cfos-tTA小鼠需要通过给予多西环素/强力霉素(Dox,通常混于食物中)来维持表达抑制状态,实验刺激前1-2天去除Dox,使得给予的特定刺激所活化的细胞产生的tTA能够结合到tetO/TRE元件上驱动tau-LacZ及其他目的基因的表达。刺激后小鼠即返回Dox抑制状态从而实现特定时间窗内的活性标记(Lewandoski. 2001)。
基于TetTag系统(含cfos-tTA转基因小鼠及病毒),Mark Mayford实验室、MIT Picower研究所的利根川进(Susumu Tonegawa)实验室、UC Davis的Brian Wiltgen实验室、日本富山大学的Kaoru Inokuchi实验室以及Alcino J. Silva实验室等研究组有一系列关于记忆印迹及事件编码神经集群的工作报道(见表5)。
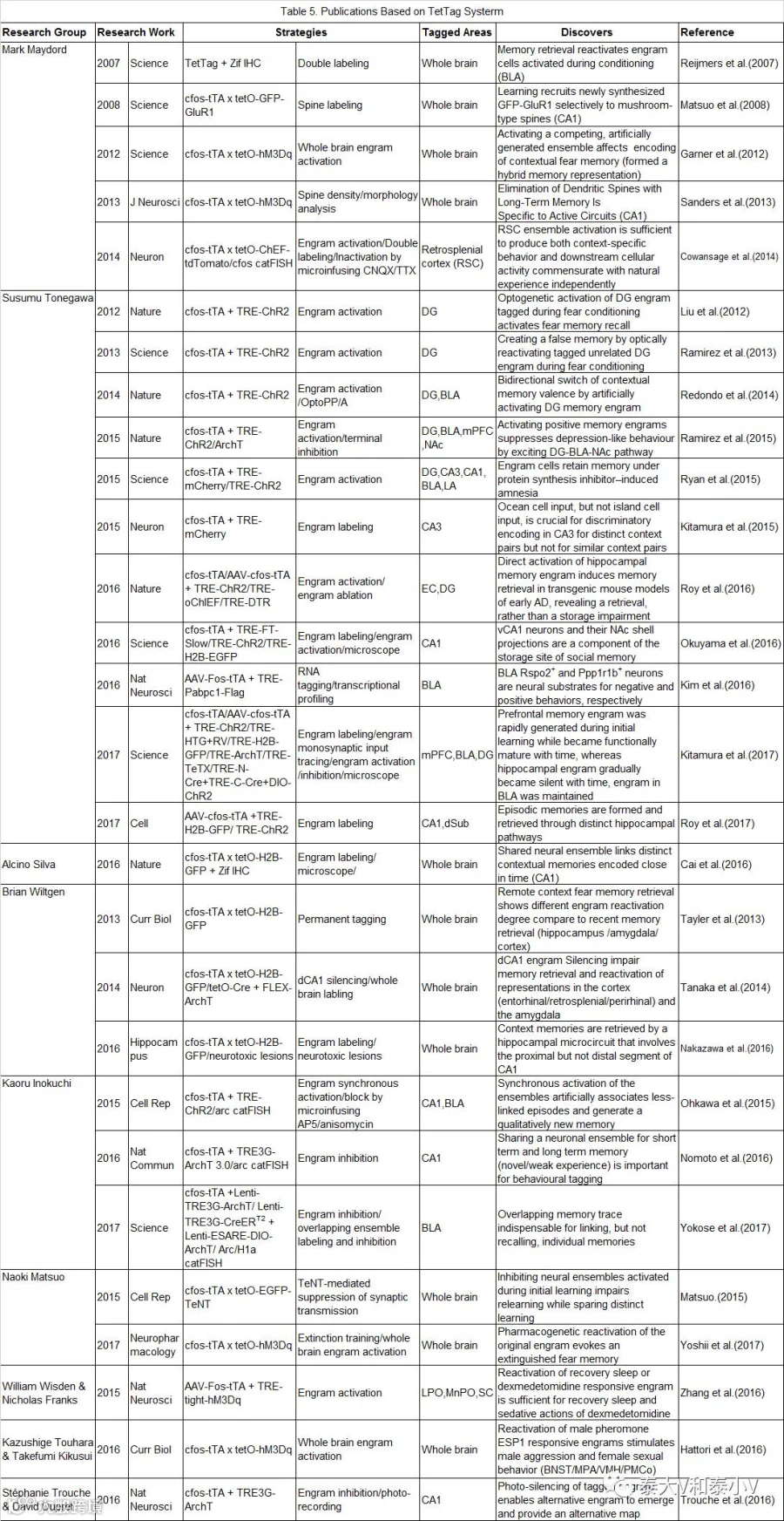
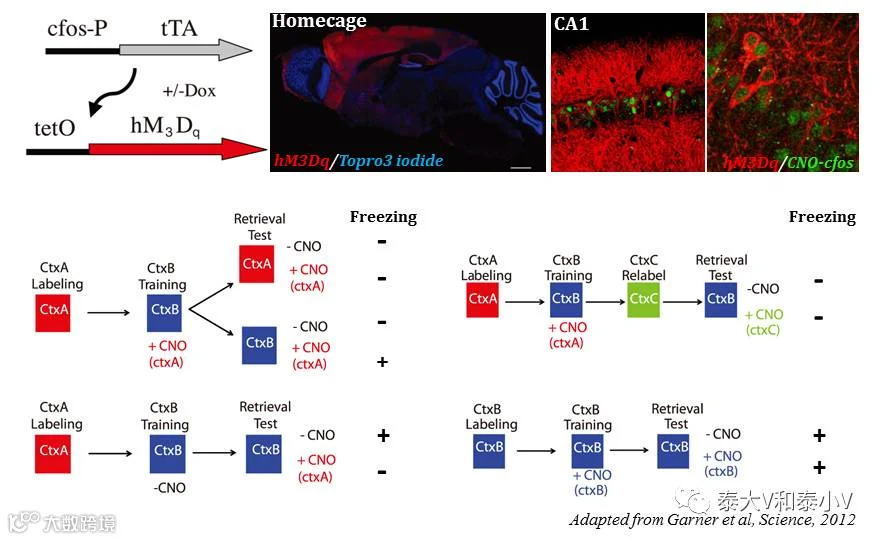
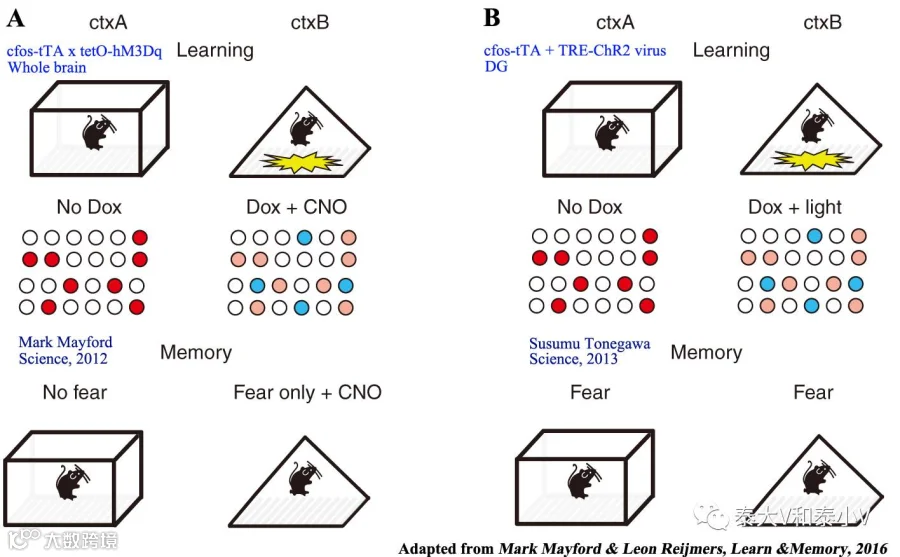
2007年Mark Mayford实验室通过TetTag工具首次证明了基底外侧杏仁核(BLA)在恐惧习得(Fear conditioning)时所激活的神经集群在记忆提取(Memory retrieval)时会被再度激活,且重激活神经元数量与恐惧记忆强度呈正相关(BLA和LA重激活分别与contextual/tone 恐惧记忆相关)(Reijmers et al. 2007)。次年他们结合cfos-tTA及tetO-GFP-GluR1小鼠在《科学》杂志再度发文,恐惧偶联学习24小时后新合成的GluR1 会被募集到成年小鼠海马CA1神经元蘑菇状树突棘(Spine)这一特定类型的Spine上去,该结果佐证了突触标签(Synaptic tagging)模型(Matsuo et al. 2008)。随着时间推进,操纵细胞活性的元件逐渐被应用流传。2012年,Mark Mayford实验室利用cfos-tTA x tetO-hM3Dq杂交小鼠通过给予外源配体CNO药理遗传激活竞争性记忆印迹致使环境恐惧记忆的形成和提取被破坏及造成记忆交联的工作发表于《科学》杂志(Garner et al. 2012)。
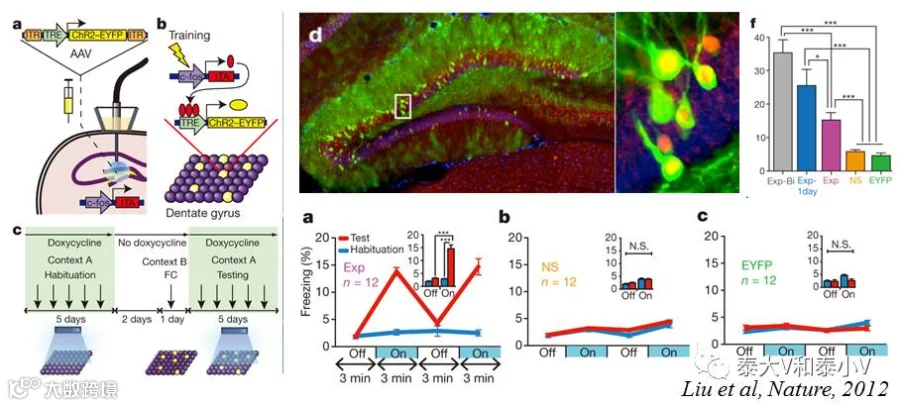
而同时期的《自然》杂志则刊载了Susumu Tonegawa实验室通过光遗传激活海马记忆印迹引起恐惧记忆召回的研究工作。他们将新构建的TRE-ChR2病毒注射到 cfos-tTA小鼠海马齿状回(DG)标记环境恐惧偶联的记忆印迹,随后通过光遗传激活标记的DG神经元引发了小鼠恐惧僵持(Freezing)表现,这为记忆印迹编码的充分性(激活足以诱导行为)提供了直接证据(Liu et al. 2012)。
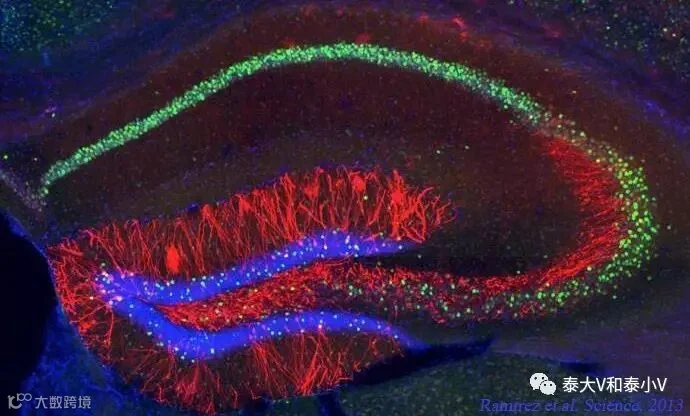
次年Science杂志以封面文章的形式刊登了Tonegawa实验室利用光遗传激活DG记忆印迹成功制造假记忆的研究工作(Ramirez etal,2013)。小鼠在环境B恐惧学习(CFC)时用光遗传激活DG里空白环境A的记忆印迹,致使小鼠放回环境A时发生记忆错乱出现僵持表现,且这一假记忆足以引发条件性位置厌恶(CPA)。同时他们证明这一记忆交错现象在海马的另一个亚区CA1是不存在的。
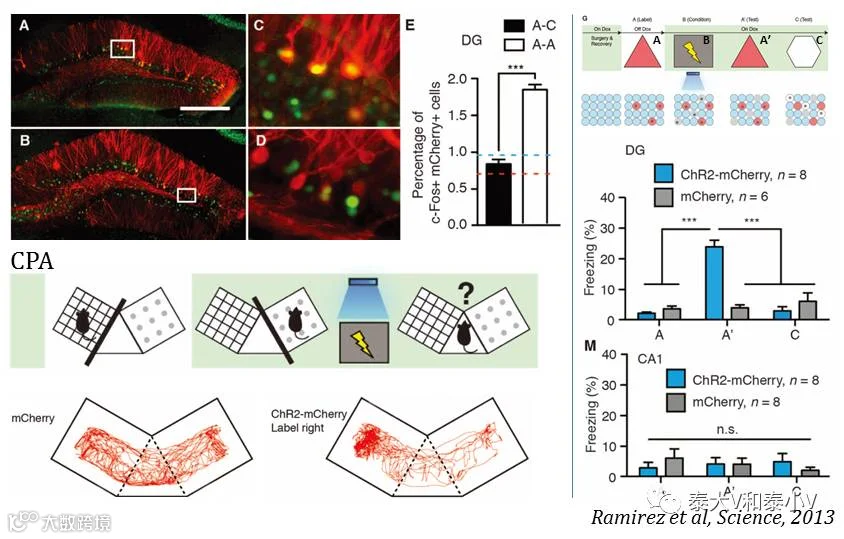
这一记忆错乱现象可能是记忆提取时的错乱,亦有可能是记忆编码时即已产生了交联,参考Josselyn募集编码的研究工作(Han et al. 2007, Park etal. 2016),这些被人工激活的DG细胞可能在条件性学习时被写入了新的恐惧偶联信息。有趣的是同样是记忆交联,前述的Mark Mayford实验室利用药理遗传人工激活全脑记忆印迹却并没有表现出原空白环境A记忆提取时的错乱(Garner et al. 2012, Mayford&Reijmer. 2016),这为记忆在神经网络中稀疏编码增添了神秘色彩。
此后Tonegawa实验室发表了一系列应用cfos-tTA小鼠及后续制作的cfos-tTA病毒研究记忆印迹的工作:
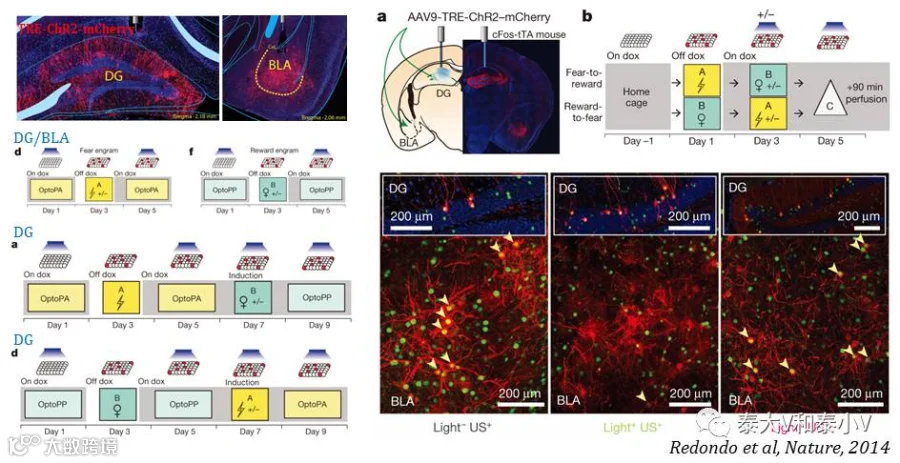
2014年在人工制造假记忆的基础上他们进一步实现了正/负情绪效价记忆(雌鼠社交经历/足底电击恐惧经历)的转换,再次证明了DG记忆印迹具有很强的可再写性,而BLA记忆印迹进行同样的人工激活程序却无法逆转。此外他们还拓展了记忆印迹环路水平的研究,首度涉及了下游脑区激活类群分析及不同脑区记忆印迹之间的关联(2 Neural ensembles),并发现DG的记忆印迹在人工交联后会支配下游不同的BLA神经集群(Redondo et al. 2014)。
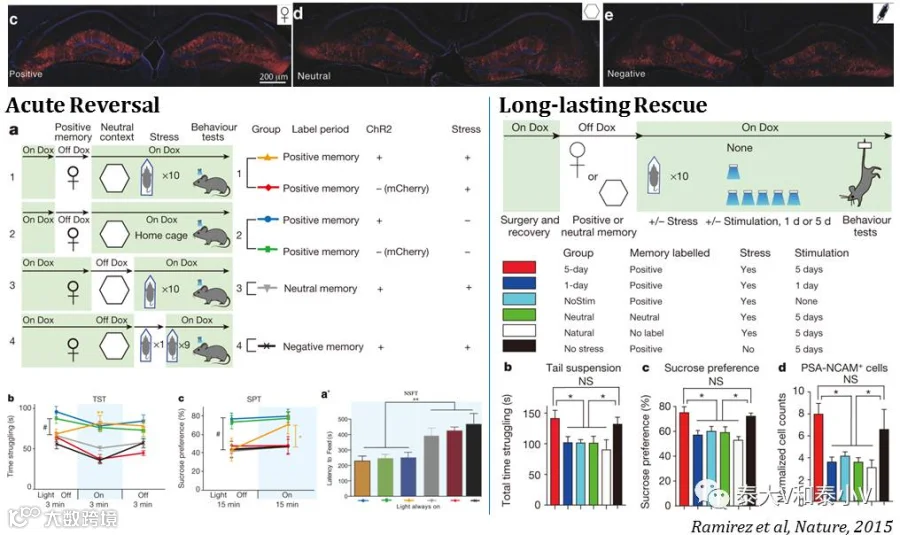
2015年则将记忆印迹引入了神经精神疾病研究:一方面是激活DG正面记忆印迹逆转慢性压力引发的抑郁(Depression)表型(Ramirez et al. 2015),另一方面是通过激活印迹找回因逆行性遗忘(Retrograde amnesia)丢失的记忆(Ryan et al. 2015)。抑郁方面的研究工作如下所示:
通过光遗传激活DG正性记忆印迹可以即时逆转连续10天2-3小时束缚应激引发的抑郁表型,包括悬尾测试(TST)挑战下放弃挣扎抵抗(Behavioral despair)、糖水偏好(SPT)缺乏(Anhedonia)及长时程新环境抑制进食(NSF)表现(Anxiety/circumvention),但并不影响小鼠在焦虑行为测试如旷场(OFT)中央区探索及高架十字迷宫(EPM)开放臂探索的表现。DG正性记忆印迹的人工激活会引发多个下游脑区的激活,随后他们即通过应用ArchT光遗传抑制投射的实验证明即时逆转效应是经由DG-BLA-NAc这一环路的激活发挥作用的,BLA而非内侧前额叶(mPFC)的正性记忆印迹及其往伏隔核(NAc)内侧壳区的输出是必要的,且这一效应受多巴胺输入调节。此外他们还证明连续5天每天上/下午各15min的DG正面印迹激活可稳定地逆转抑郁表型及恢复海马神经再生(Ramirez et al. 2015),这为抑郁治疗提供了一个别开生面的方案。
而逆行性遗忘的研究工作本质上是关于记忆巩固(Memory consolidation)和信息储存的。突触增强(Synaptic potentiation)的稳定是记忆巩固经典的细胞模型,在学习发生后的一个有限的时间窗内这一效应容易受到破坏而引发逆行性遗忘。首先他们利用脑片电生理证实了记忆印迹细胞存在与记忆巩固相关的突触增强:通过激活内嗅皮层(EC)投往DG的穿通纤维(Perforant path)末梢的同时记录TetTag工具所标记的DG环境恐惧记忆印迹细胞(mCherry+)及非印迹细胞(mCherry-),他们发现记忆印迹细胞存在更大的突触强度(AMPA/NMDA ratio),而CFC后立刻给予茴香霉素则会抹去记忆印迹的突触增强(24h后给则不会),DG印迹细胞的树突棘密度(Spine density)也表现出同样的性质。此外他们还证明了DG记忆印迹经苔状纤维(Mossy fiber)通路偏好地作用于CA3的记忆印迹,亦即印迹-印迹连接。
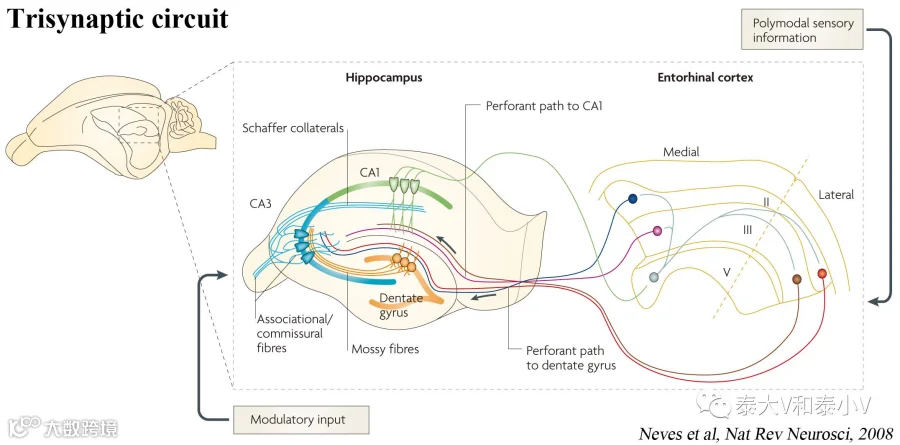
随后他们利用光遗传直接激活DG记忆印迹,出乎意料地发现逆行性遗忘小鼠的恐惧记忆(Freezing)及主动回避反应(OptoPA)都被即时找回(光依赖的短暂效应),而标记的同时如果将下游的CA1抑制,激活DG记忆印迹产生的记忆找回则消失了(上述实验中茴香霉素并未影响ChR2的合成积累)。有意思的是他们这次采用4Hz的给光程序证明CA1也是存在记忆印迹的(之前的研究采用20 Hz激活CA1标记的神经集群无效应(Liu et al. 2012)),相应的结论也与DG类似。此外他们还证明LA记忆印迹激活也可以即时找回逆行性遗忘小鼠的AFC恐惧记忆。更为有趣的是破坏记忆的重新巩固(Reconsolidation)引发的逆行性遗忘及茴香霉素破坏假记忆植入的奠基(Inception)也可被激活记忆印迹所即时恢复。最后他们通过研究DG恐惧记忆输出的下游功能脑区CA3及杏仁核的记忆印迹与DG功能连接的相关性证明了逆行性遗忘小鼠DG记忆印迹的直接激活具有和正常小鼠恐惧记忆自然召回相似的效应。逆行性遗忘很可能源于记忆巩固的失败,记忆通路(Neves et al. 2008)易感环节在记忆编码或是巩固过程中遭受破坏都可能会致使记忆的缺损(Ryan et al. 2015)。
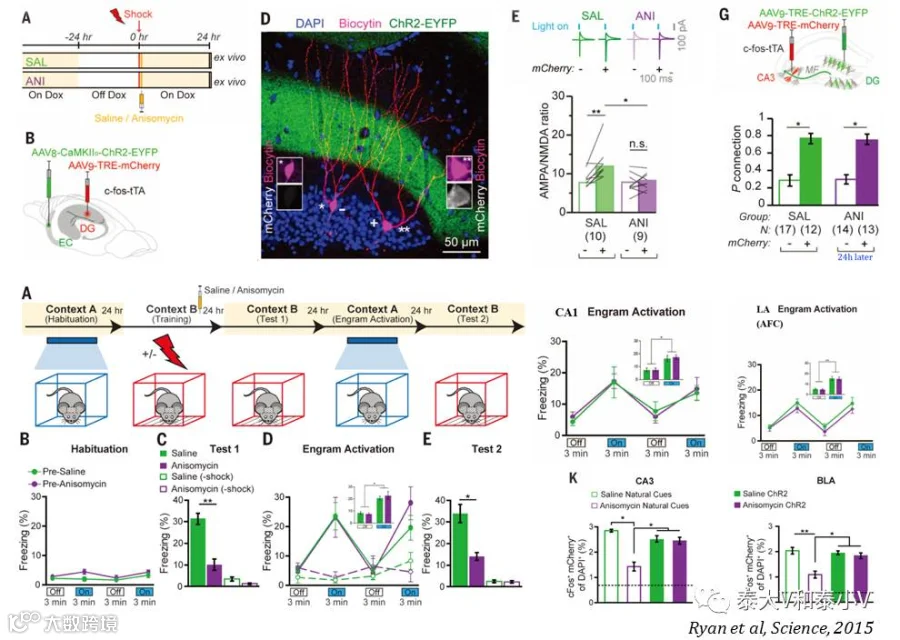
次年他们进一步用早期阿尔兹海默病(Alzheimer, AD)模型小鼠研究记忆的遗忘。AD是一种由进程性记忆衰退进而发展为认知功能缺损的神经退行性疾病(Neurodegenerative disorder),在其早期通常只表现为情景记忆(Episodic memory)的缺陷。故而他们采用7个月大的仅表现为长时程记忆(LTM, 24h)缺失而尚无Aβ严重堆积及短时程记忆(STM, 1h)缺失的AD小鼠(APP/PS1 双转基因)作为早期AD的模型,结合TetTag病毒工具他们揭示了这一病理性早期遗忘的本质在于记忆提取而非记忆储存的破坏。激活DG记忆印迹可直接取回丢失的长时程记忆,而通过光遗传(oChIEF)诱导DG记忆印迹穿通纤维突触的长时程增强(LTP)则可修复(Restore)长时程记忆及DG记忆印迹的树突棘密度,进一步利用白喉毒素毒杀(Ablation)树突棘密度修复了的DG记忆印迹则会阻断长时程记忆的恢复。此外他们还证明了早期AD小鼠在抑制性躲避(Inhibitory avoidance)及新物体探索(Novel object location)这两种不同的海马依赖的学习经历的记忆缺失也可被同样的干预方案所修复(Roy et al. 2016)。
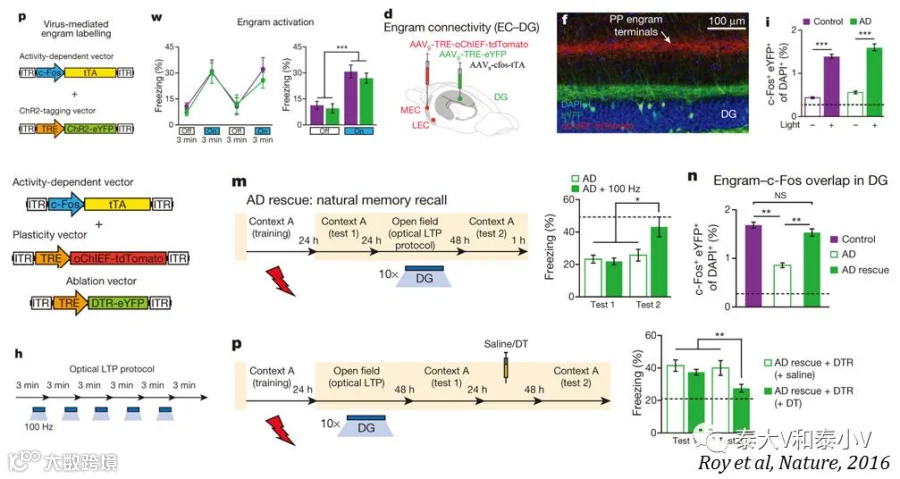
同年他们还有一篇研究腹侧海马CA1亚区及其往NAc壳区的投射储存社交记忆的工作发表于Science杂志。喜欢和陌生的小鼠社交是小鼠的天性,在经典的社交偏向实验(SDT)中,被测小鼠会花更多的时间停留在陌生小鼠的附近。在利用光遗传操纵及显微内窥镜监测鉴明vCA1→NAc Shell储存社交记忆之后,他们进一步利用TetTag工具标记及操纵了社交记忆印迹。有趣的是他们通过vCA1记忆印迹激活奠基成功地将正性(可卡因刺激)/负性(足底电击)的经历植入到与小鼠A的社交记忆里,引发了实验测试小鼠对小鼠A的喜好/厌恶情绪,这证明vCA1也具有很强的可写入性(Okuyama et al. 2016)。
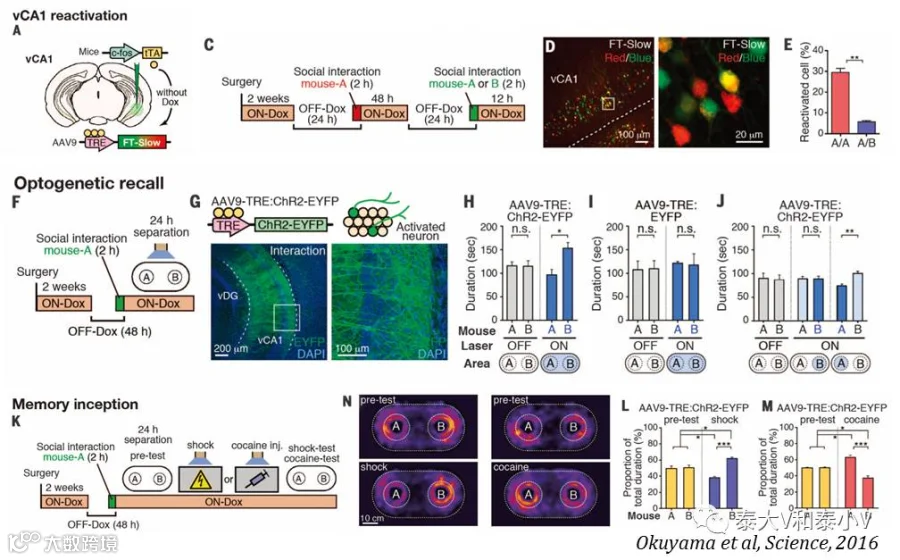
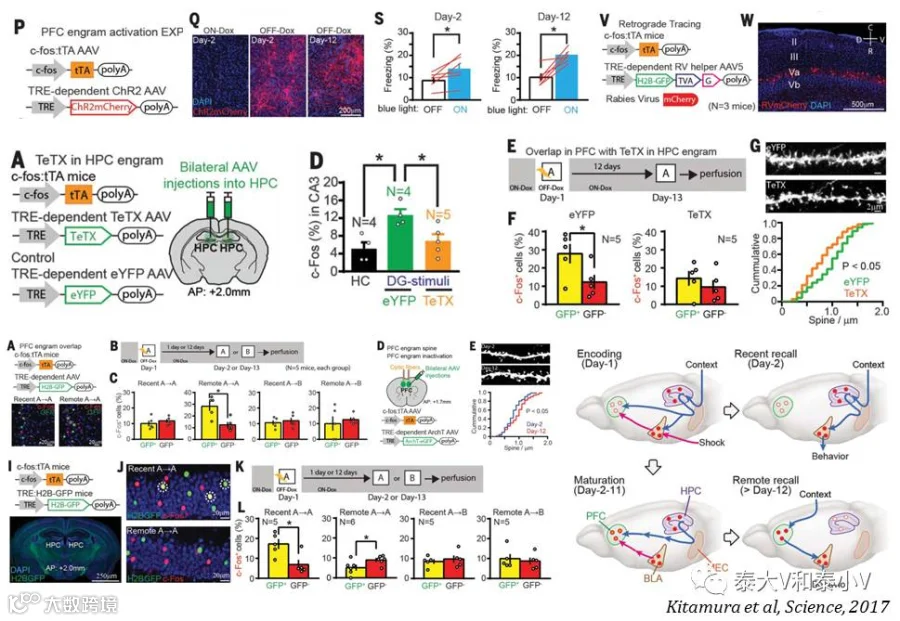
2017年他们将视线转向了情景记忆在皮层永久储存背后的记忆巩固系统(Kitamura et al. 2017)。结合活动依赖工具标记记忆印迹、环路追踪、系统操纵及显微内窥镜监测的研究,他们证明皮层所储存的长期记忆并非完全是由形成和储存短期记忆的海马转移而来,内侧前额叶(mPFC)在最初CFC时即已通过海马-内嗅皮层网络及BLA的信息输入产生了记忆印迹,但是这些早期的皮层记忆印迹除了光遗传直接激活会引发小鼠僵持表现之外并不编码环境偶联信息(来自TetTag标记+cfos IHC及显微内窥镜监测的证据),随着时间推移,在海马记忆印迹的帮助下他们储存长期记忆的功能逐渐成熟,而海马的记忆印迹逐渐沉默,但与功能输出相关的下游BLA的印迹则一直保持活跃,由此他们重构了一个记忆系统巩固的新模型。mPFC在恐惧记忆初始时即已产生行为表现相关的神经集群的结论与Karl Deisseroth实验室所证明的mPFC存在情绪编码神经集群的研究(见后文)(Ye et al. 2016)相符,但这一研究有趣的地方在于恐惧记忆长期储存时将情景环境偶联的信息流从海马等结构写入到了前述的mPFC响应足底电击(Shock-responding)的神经集群,突触密度增加进而产生了存储长期记忆的成熟印迹。有意思的是早先的Nature杂志刊登了一篇东京大学Haruo Kasai实验室利用新开发的突触光探针AS-PaRac1标记及光反应擦除运动皮层突触记忆印迹的文章(Hayashi-Takagi et al. 2016),力证信息储存于树突棘-突触这一丰富的结构单位的可塑性变化(Synaptic remodelling)之中。
此外Alcino J. Silva实验室及Kaoru Inokuchi实验室各有一项研究偶联记忆通过共用印迹以关联的出色的工作发表(Cai et al. 2016, Yokose etal. 2017),后者更是结合Tet-off及CreER系统标记及操纵了共用的神经集群(Overlapping memory trace)。另2014年Mark Mayford实验室发表于Neuron杂志的文章证明压后皮质(RSC)存在位置反应印迹细胞,也可以写入恐惧偶联信息,且在随后的人工激活中引发僵持表现及造成恐惧偶联的消退(Extinction)(Cowansage et al. 2014)。
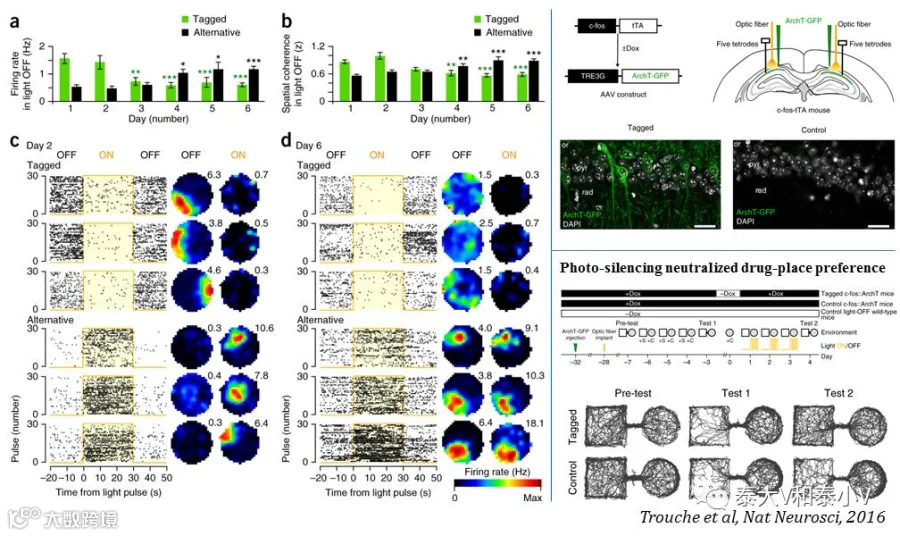
还有一项来自牛津大学MRC的David Dupret实验室的有意思的工作值得一提:他们利用TRE3G-ArchT病毒标记cfos-tTA小鼠海马CA1特定环境的记忆印迹,随后放回该环境并在光抑制印迹的情况下进行多通道记录,发现CA1产生了编码这一特定环境的新的印迹,且原本与环境偶联的可卡因药物的记忆也被消除了,大脑对这一药物偶联环境产生了新的记忆图谱(Trouche et al. 2016)。
虽然TetTag被大量用于记忆印迹等方面的研究,但TetTag存在转基因动物的通病:其表达可能与内源表达模式不一致。此外Tet-off系统需要1-2天清除体内Dox及刺激后返回Dox抑制状态,使得系统开放的时间窗较大致使背景泄漏偏高(Guenthner et al. 2013)。不过基于Tet的系统具有快速高水平表达目的基因的特质(诱导次日即可进行实验测试),这对于研究诸如近期记忆提取等短时程变化具有重要意义。
TRAP
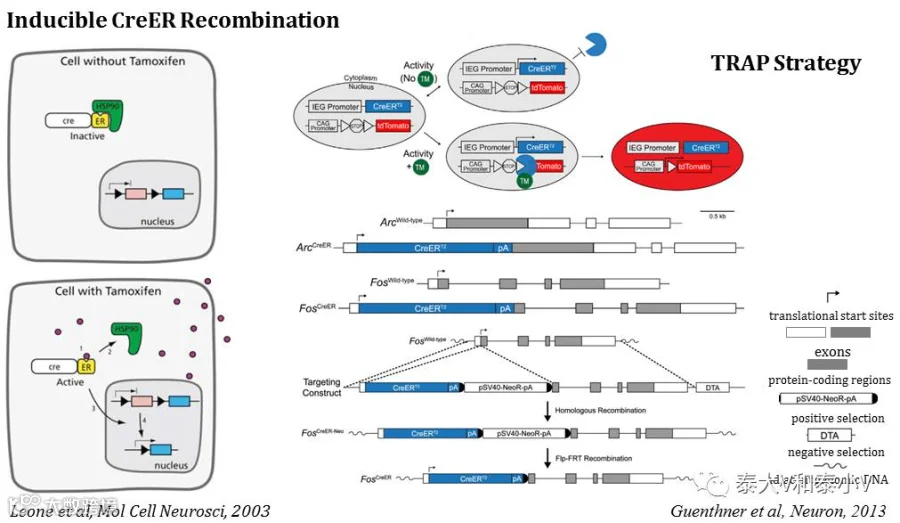
活性类群靶向重组(TRAP)工具是斯坦福大学的骆利群(Liqun Luo)教授在2013年发表cfos-CreERT2和Arc-CreERT2 基因敲入(KI)小鼠(Guenthner et al. 2013)时所提出的概念,这里我们将哥伦比亚大学René Hen教授2014年发表的Arc-CreERT2转基因(BAC-Tg)小鼠(Denny et al. 2014)也收入其中。TRAP工具的核心在于通过他莫昔芬(Tamoxifen)及其代谢产物4-OHT诱导CreER入核重组实现条件性基因表达:CreERT2是P1噬菌体重组酶Cre与人雌激素受体配体结合域(LBD)三点突变体(G400V/M543A/ L544A突变,不再与其天然配体17β-雌二醇结合)的融合蛋白。自然情况下,细胞生成的CreERT2被热激蛋白HSP90结合而锁定在细胞质,只有在给予人工合成的配体Tamoxifen/4-OHT的情况下,4-OHT结合到ERT2-LBD使得HSP90解离,CreERT2进而转位到细胞核特异性识别LoxP位点从而介导重组的发生,使得基于同向LoxP的LSL元件的Stop序列切除或是基于反向LoxP的DIO/FLEX衔接颠倒的目的基因元件倒转,从而实现目的基因的表达(Feil et al. 1997, Leone et al. 2003, Matsuda & Cepko. 2007)。
cfos/Arc-CreERT2(KI) 与 Ai14(R26-LSL-tdTomato:deltaNeo)杂交产生的荧光报告小鼠的一系列测试表明:FosTRAP几乎不存在系统泄漏、重组依赖于特定的刺激输入(见经典的胡须拔除实验)、拥有比TetTag系统更小的标记时间窗(见视觉皮层V1光刺激反应实验,4-OHT的诱导时间窗小于12小时),且具有高达70%的特异性和30-40%的外显率(见蜗背侧核对高频/低频声音刺激的标记实验);
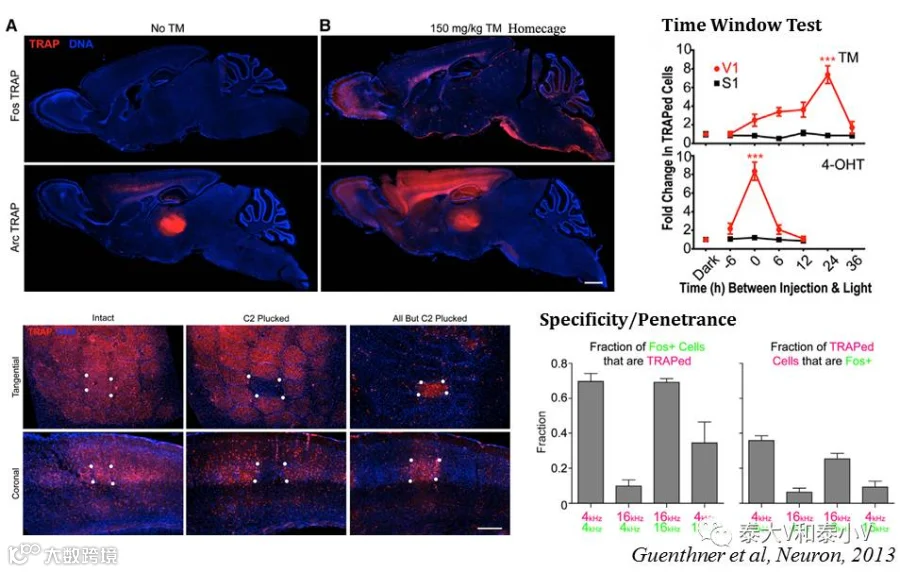
ArcTRAP具有比FosTRAP更高的重组效率,但在部分脑区的背景泄漏很高。此外出于CreERT2表达水平可能很低,重组效率不足的担忧,骆利群实验室在开发TRAP工具时直接将CreERT2敲入到小鼠即早基因cfos/Arc启动子的后面,这一策略所产生的无效等位基因破坏了内源Fos的表达,而之前有报道Fos+/-杂合子小鼠存在单倍剂量不足神经表型,诸多原因致使TRAP的应用受限(Sakurai et al. 2016),后续应用的报道不多(见表6)。
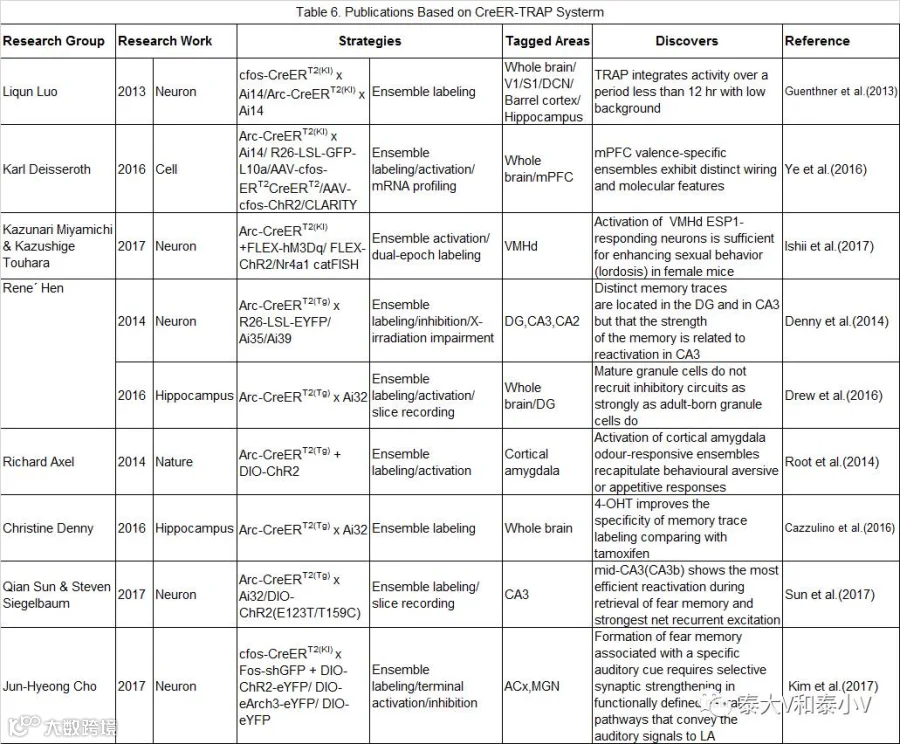
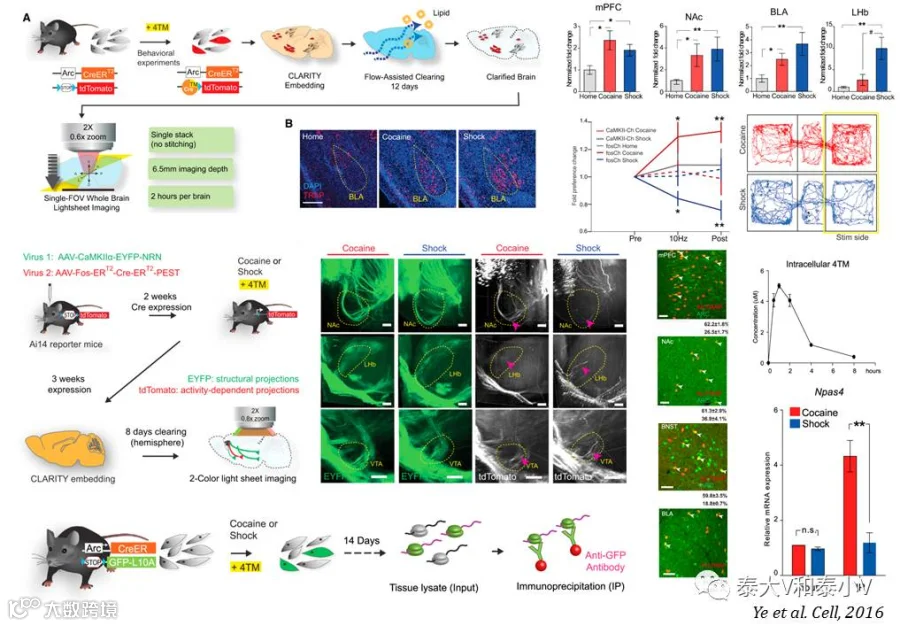
但是除了前文所述的TetTag系统标记之外,CreER-TRAP诱导重组的方案是目前仅有的可以进行啮齿类全脑印迹标记的策略,对于高通量对比筛选具有重要意义。斯坦福大学Karl Deisseroth实验室即有一项基于ArcTRAP(KI)的研究报道:借助CLARITY透明化及高通量对比分析可卡因/足底电击处理后的Arc-CreERT2(KI)x Ai14杂交报告小鼠全脑正/负面刺激响应的神经集群,他们绘制出了相应的全脑激活图谱并分选出了对正/负面刺激具有差异性响应的脑区,外侧缰核(LHb)。随后他们通过Ai14小鼠注射新构建的活动依赖病毒AAV-cfos-ERT2-CreERT2-PEST进行神经集群荧光标记及通过Arc-CreERT2(KI)x R26-LSL-GFP-L10a杂交小鼠进行核糖体分型(Ribosome profiling)分别解析了PFC不同情绪效价响应的神经集群存在的环路基础(Wiring features)及分子基础(Molecular features),此外他们使用AAV-cfos-ChR2-PEST病毒通过光遗传激活PFC特定的神经集群直接诱发了相应的行为表型,证明了PFC存在情绪的神经编码(Ye et al. 2016)。受所用工具本身的限制,这一大数据研究的部分细节还有待优化。据悉骆利群实验室正在开发TRAP 2.0,势必会有巨大的改进突破。
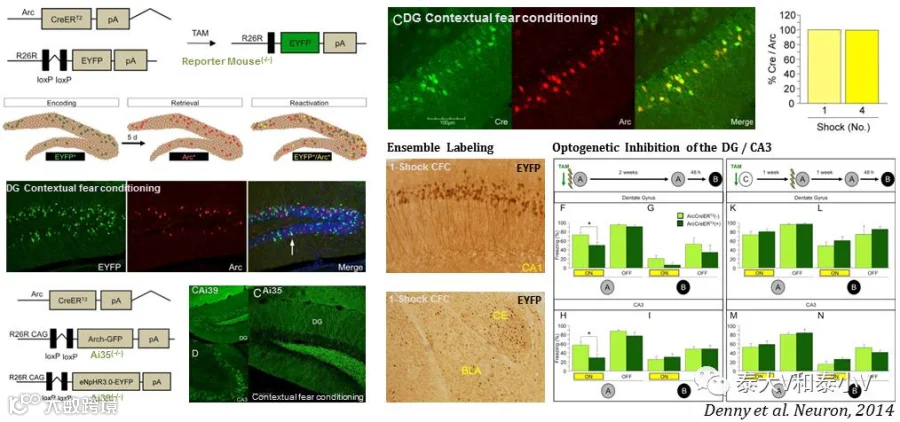
René Hen实验室所开发的 Arc-CreERT2(Tg)BAC转基因小鼠最早被用于海马记忆印迹标记及抑制:通过Arc-CreERT2(Tg) xR26-LSL-EYFP杂交报告小鼠标记恐惧环境记忆编码(Memory encoding)印迹及Arc/cfos免疫组织化学染色(IHC)表征恐惧记忆提取(Memory expression)所激活的印迹相结合的方法,他们的研究表明小鼠海马DG及CA3记忆印迹随着时间推移记忆泛化(Generalization)而沉默(亦即长期记忆储存的转移)。另他们通过光遗传抑制Arc-CreERT2(Tg) xAi35(R26-LSL-Arch-GFP)/ Ai39(R26-LSL-eNpHR3.0-EYFP)杂交小鼠DG/CA3的记忆印迹发现相应记忆的提取被削弱,但抑制CA2亚区则不会引发相似的效应,印迹会定位于核团具体的功能亚区(Denny et al. 2014)。
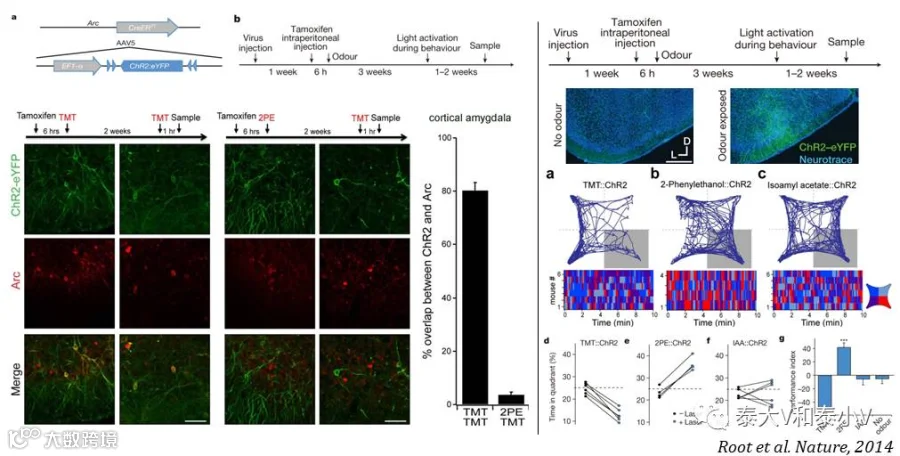
Arc-CreERT2(Tg) 转基因小鼠另一有意思的应用是同年发表于《自然》杂志的与哥伦比亚大学Richard Axel 实验室合作的关于气味引发内在行为相关的皮质杏仁核(Cortical amygdala)神经集群的研究工作(Root et al. 2014)。他们首先利用光遗传抑制嗅球-皮质杏仁核投射的实验证明这两个脑区在TMT/2PE两种气味分子所分别引发的厌恶回避/喜好吸引行为中是必要的,之后应用活动依赖小鼠标记并激活响应两种气味分子的皮质杏仁核神经集群复现了两种气味所引发的行为表现,为皮质杏仁核存在气味驱动先天行为编码提供了直接证据。
CANE
活化神经集群捕获(CANE)工具是杜克大学王帆(Fan Wang)实验室所开发的一套全新的基于可降解受体dsTVA的活性依赖标记系统。该系统借助嗜神经病毒应用时所采用的衣壳包裹伪装病毒-靶点细胞表达受体两者特异结合的方案,首先是制作了一个基因敲入小鼠品系cfos-2A-dsTVA,使得被刺激活化的细胞在cfos的驱动下表达易于降解的鸟肉瘤/白血病病毒包膜蛋白(EnvA)的特异受体TVA,生成的TVA模拟了内源cfos的代谢,随后会嵌入活化细胞的膜表面提供靶标并在较短时间内被降解;此时将EnvA包裹伪装的狂犬病毒(SAD-ΔG-RV-mCherry)或是慢病毒(LV-hSyn-Cre)及时注入到目标脑区,EnvA-TVA结合使得刺激活化的细胞感染携带了目的基因结构的RV/LV从而被捕获标记(Sakurai et al.2016)。这一可降解受体方案使得工具的写入性(Versatility)得到了极大的扩展,每次标记只要注入含有不同的目的基因结构的EnvA包裹伪装的病毒(XFP/Cre/Flp/Dre)即可实现同一脑区多个类群的标记及干预。这是活动依赖工具开发的一次重大突破。
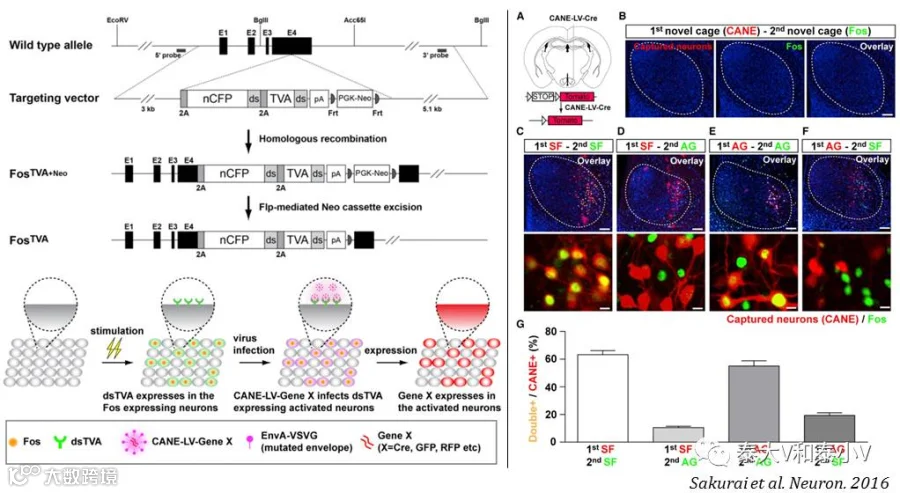
为检验CANE的特异性及效能,王帆实验室应用雄性啮齿类经典的入侵者实验(Resident-intruder assay)中社交冲突双方的攻击行为和恐惧都会激活下丘脑这一与攻击(Aggression)、社交恐惧(Social fear)、求偶及性行为(Mating)等先天行为有关的脑区的特点,研究发现腹内侧下丘脑腹外侧区(VMHvl)这一亚区存在两群相互分离但交错分布(Intermingle)的神经集群分别响应攻击(AGNs)及社交恐惧(SFNs)。CANE标记具有60%以上的特异性及50%左右的外显率,而随后光遗传激活及破伤风毒素(TeLc)突触传递抑制VMHvl SFNs的实验则进一步证实了这一神经集群编码社交恐惧的充要性。另借助顺行标记/逆行跨单突触追踪这一神经集群的输入/输出,他们进一步揭示了VMHvl SFNs存在高度离散但相互连结(Recurrent)的环路结构(Circuit architecture)。
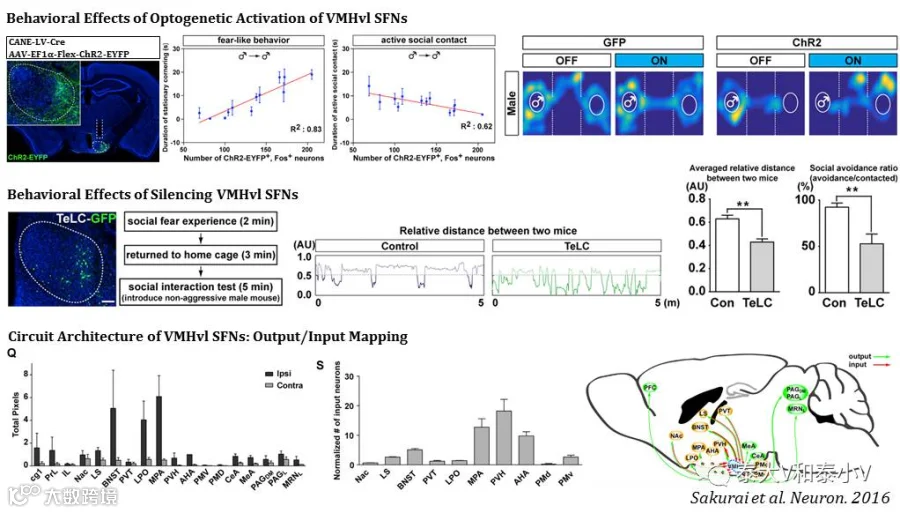
此外他们还对比分析了CANE分选标记的功能类群与经典的分子标记类群的共定位情况,发现雌激素受体1(Esr1+)/类固醇生成因子-1(SF1+)并不是理想的AGNs/SFNs的分子标签。这一结果亦表明大脑功能编码的复杂性,而利用活动依赖工具进行功能解析具有重大的应用前景。相比于TetTag和TRAP工具,CANE具有诸多优点(见表7),但由于需要在刺激1-2小时后及时将包裹伪装的病毒注入目标脑区,动物麻醉及手术损伤都可能会引发无关的背景标记,提前置入给药导管如无线光流控系统(Wireless optofluidic systems)(Jeong et al, 2015)或许是一个可行的解决方案。
E-SARE & RAM
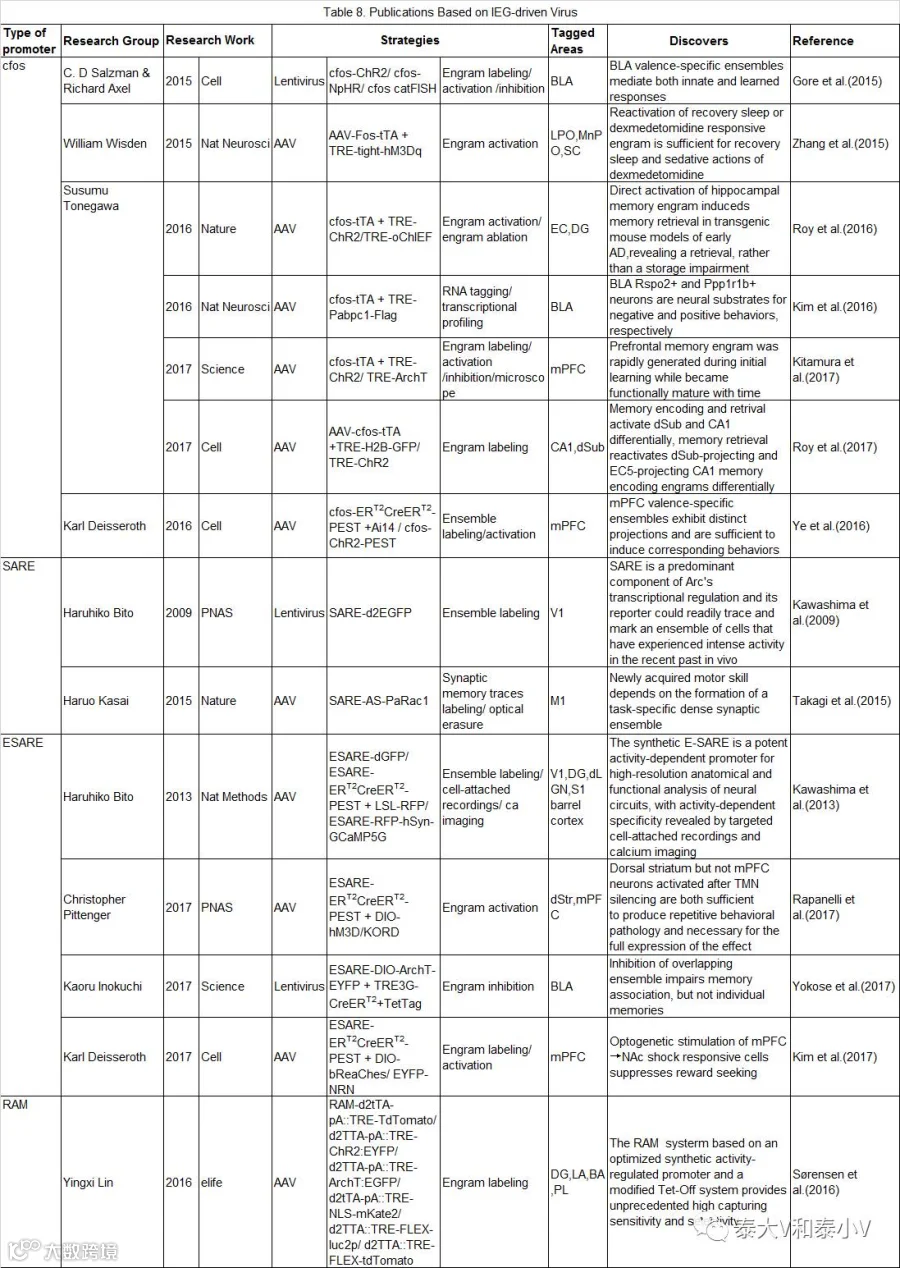
相比于实验动物工具,病毒工具具有更大的灵活性和适用性(Flexibility)。哥伦比亚大学的C. D Salzman/ Richard Axel实验室、Susumu Tonegawa实验室、Karl Deisseroth实验室及英国帝国理工学院的Nicholas Franks /WilliamWisden实验室都各自开发了不同的cfos病毒工具(见表8)。
2015年C. D Salzman/ Richard Axel实验室通过fos-ChR2/NpHR慢病毒工具揭示BLA存在正性刺激(尼古丁刺激)/负性刺激(足底电击)响应的神经集群介导相应的先天及习得性反应的研究工作发表于《细胞》杂志。结合病毒活性标记和cfos IHC或是cfos catFISH的双事件标记(Dual-epoch labeling)的结果都表明BLA存在两群交织错落的神经集群分别响应尼古丁刺激和足底电击,光遗传激活这两个神经集群会引发相应的内在反应如心率、呼吸速率增减及僵持表现,随后他们进一步证明这种激活效应可以直接充当巴普洛夫联合学习(Pavlovian conditioning)的非条件性刺激(US),驱动喜好性及厌恶性学习如听觉恐惧偶联(Auditory fear conditioning)、气味偶联(Olfactory conditioning)及操作性学习(Instrumental conditioning)。光遗传抑制这两个神经集群则会破坏偶联的条件性刺激(CS)所引发的条件反射。另激活响应恐惧偶联的CS的神经集群也可以引发僵持表现,学习将CS及US响应的神经集群联系在了一起(Gore et al. 2015)。
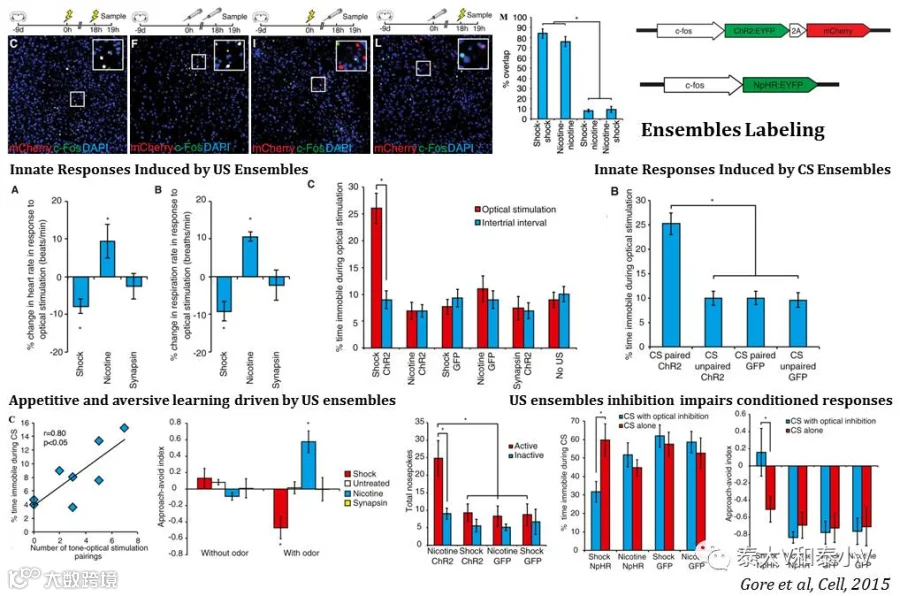
虽然文章提供的数据显示Lenti-cfos病毒的特异性及外显率能分别达到80%和50%,但由于缺乏表达时间窗调控元件,病毒表达背景存疑。
Fos mini启动子经过多年的开发一直存在效率不足且信噪不佳的问题(Sørensen et al. 2016),人工改造合成更为强劲而精良的活性启动子的工作从未止步。早在2009年日本东京大学的Haruhiko Bito教授就发现并解析了Arc转录调控的重要元件SARE并尝试开发了SARE-Arc mini启动的慢病毒工具(Kawashima et al. 2009),2013年他们实验室进一步推出了人工改造的强力活性启动子E-SARE(Kawashima et al. 2013)。E-SARE(975 bp)由5个串联重复的SARE与含TATA盒子的Arc mini融合而成。他们首先制作了一个E-SARE启动表达不稳定版GFP(dGFP)的AAV病毒,在体测试表明10min光刺激及1.5h新环境探索可以分别在视觉皮层和DG诱导显著的GFP表达,工具标记结合Arc/cfos IHC 的结果显示标记的特异性大于60%且外显率高于50%(需要注意的是,与前述的通过两次刺激诱导进行检测的方法不同,这里是同一次刺激的诱导结果)。随后他们将E-SARE-dGFP病毒注入桶状皮层这一响应单根胡须刺激的经典脑区,结合喷气(Air puff)刺激胡须及在体单细胞电生理记录(Cell-attachedrecordings),成功地证明工具标记的细胞显著地响应外加刺激(Evoked spikes)(而非自发活动(Spontaneous firing)),为工具特异性提供了在体电生理证据。此外他们还开发了E-SARE-ERT2CreERT2-PEST诱导重组系统,实现了小鼠外侧膝状体核(LGN)光激活的同侧眼神经类群向视皮层V1神经投射的永久标记。另外通过使用E-SARE-RFP-hSyn-GCaMP5G感染V1,结合双光子钙成像检测特定方向视觉刺激响应的神经集群的标记,发现钙信号强度与工具驱动的RFP表达显著相关。基于上述实验E-SARE成为了目前特异性验证最为全面的活动依赖工具。
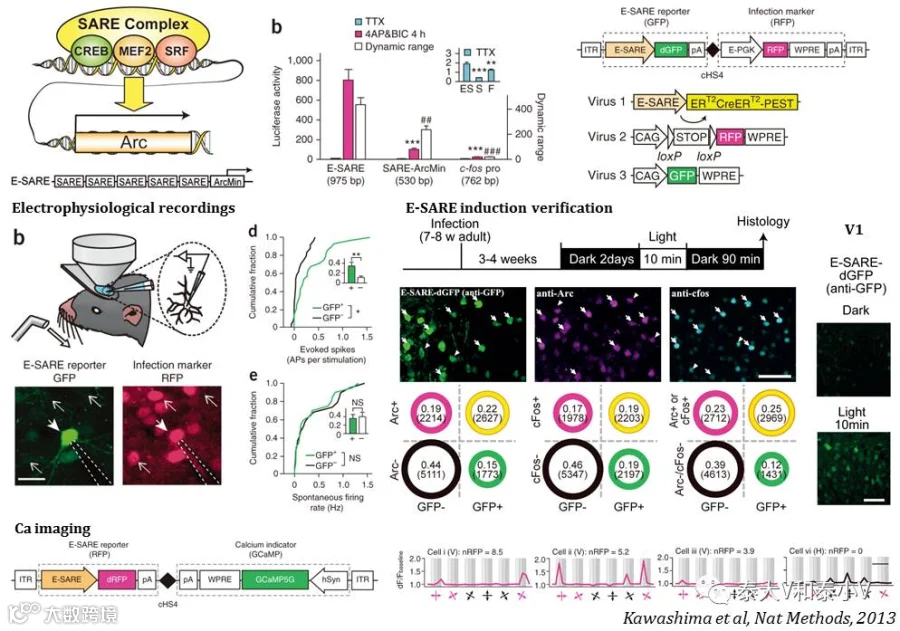
2016年MIT的林映晞(Yingxi Lin)实验室则改造合成了基于Fos mini的RAM启动子。为获取更加高效、通用及强扩展性的活性启动子,他们首先进行了多达11830个已鉴明的神经元活性调控增强子的筛查,发现AP-1位点(TGANTCA)是最为富集的一段基序。考虑到转录因子与增强子组合调控基因表达的特点,他们进而将神经元活性依赖基因Npas4的结合基序NRE(TCGTG)加载到AP-1上去,随后将双碱基重叠(10mer)的NRE/AP-1核心基序(TCGTGANTCA)插入到CME这一与转录激活有关的重要序列,形成了一个24bp的增强子模块。而4个增强子模块串联重复再连接Fos mini即生成了199bp的人工活性启动子PRAM。荧光素酶活性的测试的结果表明PRAM具有比E-SARE更高的信噪比(Sørensen et al. 2016)。
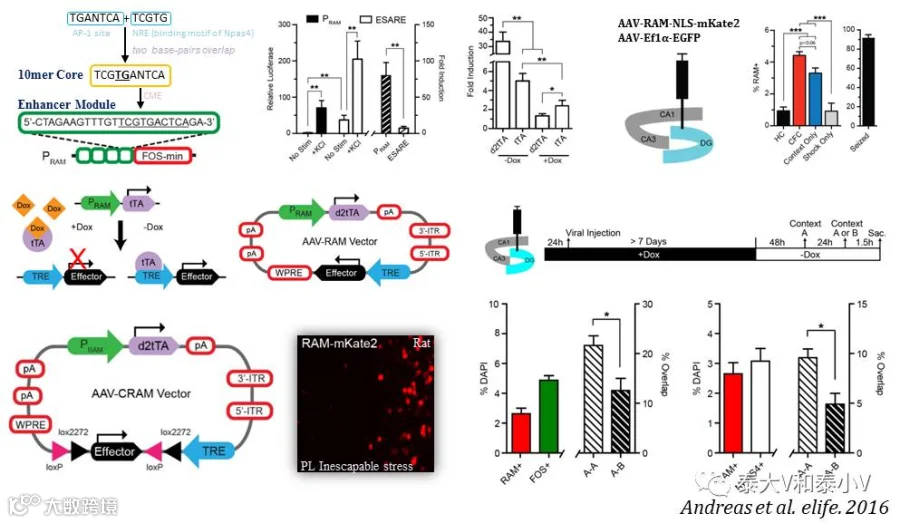
PRAM启动不稳定版d2tTA及TRE驱动效应蛋白(tdTomato/NLS-mKate2/ChR2-EYFP/ArchT-EGFP)表达的双启动子系统置于同一个AAV载体(包装容量4.9kb的AAV还剩有高达1.8kb的空间可用于加载效应蛋白)即合成了强活性标记的RAM系统。随后他们在海马DG亚区、杏仁核LA/BA及CeA亚区分别进行了在体标记测试,并尝试将RAM系统应用于大鼠及果蝇,另还开发了Cre依赖的CRAM系统(RAM-FLEX-tdTomato)用于细胞类型或是投射特异(结合可逆向运输的CAV-Cre使用)的活化神经集群的标记。由于缺乏理想的特异性验证试验及标记类群功能操纵的数据,RAM工具的在体性能还需根据具体实验进一步验证。
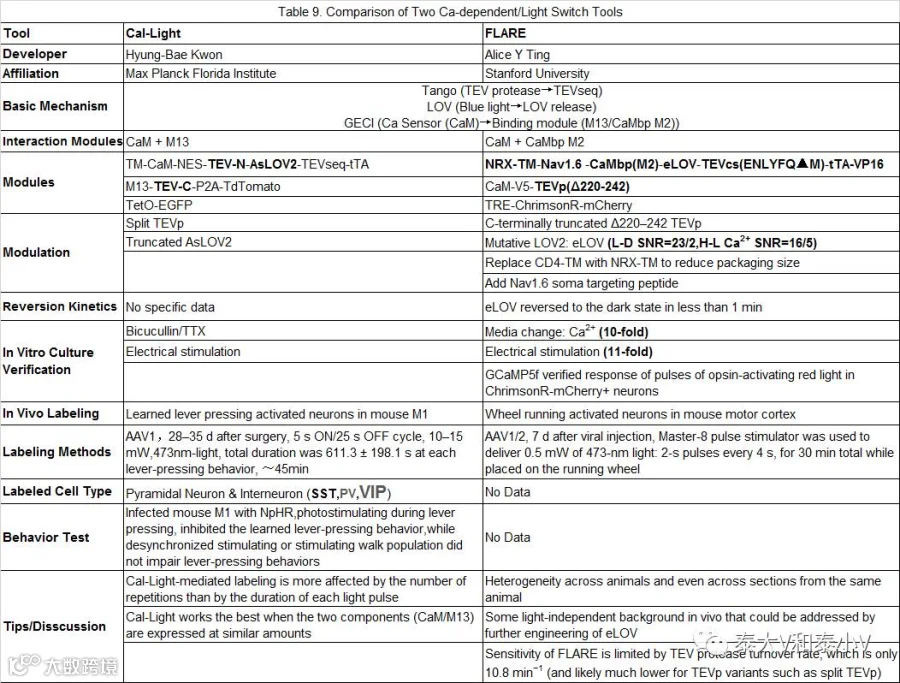
Cal-Light & FLARE
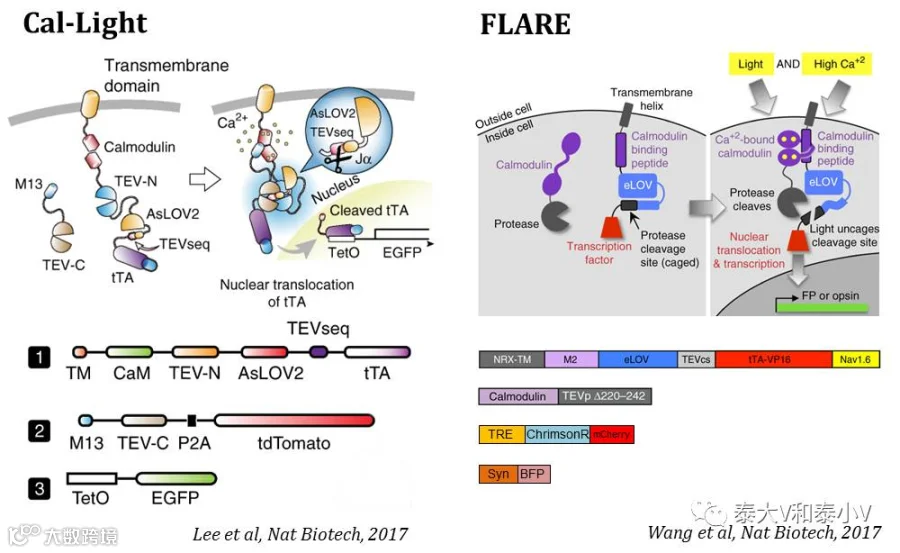
虽然IEG表达作为神经元激活的一种间接表征被广泛应用,但是对于即时标记和模拟信息流高速处理的大脑而言,其时间分辨率及泛表征灵敏性是远不足够的,开发基于即时的细胞神经化学变化的活性工具一直以来是亟待突破的方向。在历经了检测神经调节(Neuromodulation)作用的Tango(Barnea et al. 2008;Kroeze et al. 2015)及iTango2(Lee et al. 2017)系统开发等一系列出色的探索工作的基础上,终于,2017年6月26日《自然·生物技术》杂志同期在线发表了两篇关于Ca2+内流依赖、光控表达的活动依赖工具的文章,分别介绍了马普弗罗里达神经科学研究所(MPFI)Hyung-Bae Kwon实验室所开发的Cal-Light(Lee et al. 2017)和斯坦福大学Alice Y Ting实验室所开发的FLARE(Wang et al. 2017)。Cal-Light和FLARE有着近似的工作原理,他们都是基于蛋白互作系统Tango的模块式基因表达组构、基因编辑钙指示剂(GECIs)的钙检测以及燕麦向光素光氧电压结构域(LOVs)的光反应特性。
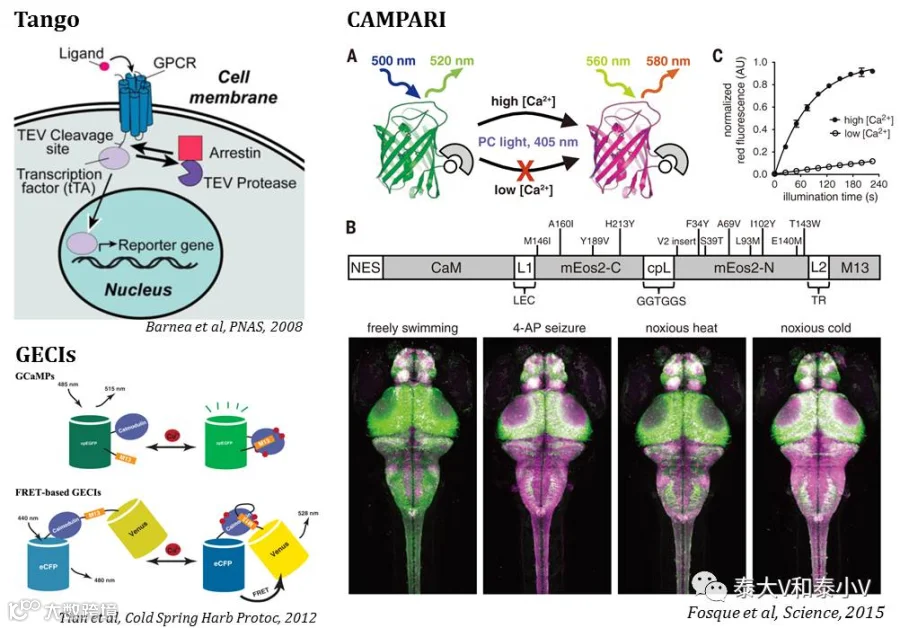
Tango起源于Notch信号转导酶切片段转位入核调控转录的机制以及烟草蚀刻病毒蛋白酶体(TEV Protease,TEVp)分裂片段互补监测蛋白互作的方案(Barnea et al. 2008)。将转录因子(如tTA)经由含TEVp酶切位点的序列连接在膜表面的GPCR, RTK或是类固醇激素受体胞内段从而栓系在细胞膜上,当配体结合于上述受体激活信号转导时,以GPCR为例,磷酸化的受体会募集胞内的β-arrestin(抑制G蛋白进一步激活的阻遏蛋白,现融合了TEVp)靠近GPCR,促使TEVp识别并酶切掉连接序列,tTA进而从膜上释放下来,入核启动目的基因表达。Cal-Light/FLARE在Tango模块式组构调控基因表达的基础上引入了GECIs来检测胞内钙离子以实现细胞活化水平的探测:GECIs包括含有Ca2+结合结构域的钙调蛋白(Calmodulin, CaM)、钙调蛋白结合肽M13及融合表达的嵌合荧光蛋白(Tian et al. 2012)。Ca2+结合于CaM会使之构象发生改变, M13进而能够与CaM结合引发嵌合荧光蛋白构象变化而使之发光(Tian et al. 2012)。于是他们将CaM和M13或M2替换了Tango系统中的GPCR和β-arrestin,将其中的一个模块与跨膜结构域(TM)及TEVp酶切序列一起把tTA栓系在膜上,而另一模块则融合了TEVp,利用胞内Ca2+作为激活整个系统的配体从而实现了Ca2+依赖的基因表达。紧接着为了实现系统对特定刺激的标记,他们进一步引入了光控元件LOV掩藏TEVp酶切位点,使得只有在蓝光照射引发LOV构象变化的情况下,TEVp酶切位点才会暴露出来,促成后续的酶切反应,由此搭载了非常强的时间精度的控制元件,从而开发出了光控Ca2+依赖的新工具Cal-Light和FLARE。值得一提的是,在Cal-Light和FLARE开发出来之前,珍妮莉娅法姆研究学院(HHMI, Janelia Farm ResearchCampus)的Eric R. Schreiter实验室开发了具有很强参考价值(Inspiring)的基于Ca2+实时检测的钙调光激活比率集成器(CAMPARI)永久性活性快门(Snapshot)技术。同样是借助GECIs检测胞内钙浓度,CAMPARI利用紫外照射分裂片段互补后呈绿色荧光的Eos光可变荧光蛋白,使之转变成红色,实现了斑马鱼幼虫、果蝇全身及小鼠视皮层相应神经集群的永久标记(Fosque et al. 2015)。但是由于诱导转化的紫外照射存在损伤作用以及无法将操纵元件载入激活的神经集群,CAMPARI的应用受到一定的限制。
Cal-Light
为实现光控钙依赖活性标记,Hyung-Bae Kwon实验室测试了超过300种不同的构建方案,最终采用了Tango的组构式基因表达方案,并将TEVp分裂成N末端和C末端两个片段分别置入到CaM和M13所在的两个模块。只有在胞内Ca2+浓度升高的情况下,M13才会与CaM结合使得TEV的N末端与C末端靠近从而获得蛋白酶体活性,此时给予蓝光照射释放出TEVp切割位点,tTA随即被切割下来入核激活TRE,启动目的基因表达。化学刺激(Bicuculline)海马培养神经元及电刺激皮层培养脑片的测试都表明Cal-Light调控的报告基因表达存在很强的刺激及光共同依赖(Coincident detector)的特性,光在刺激之前或之后给都无法启动有效的基因表达。
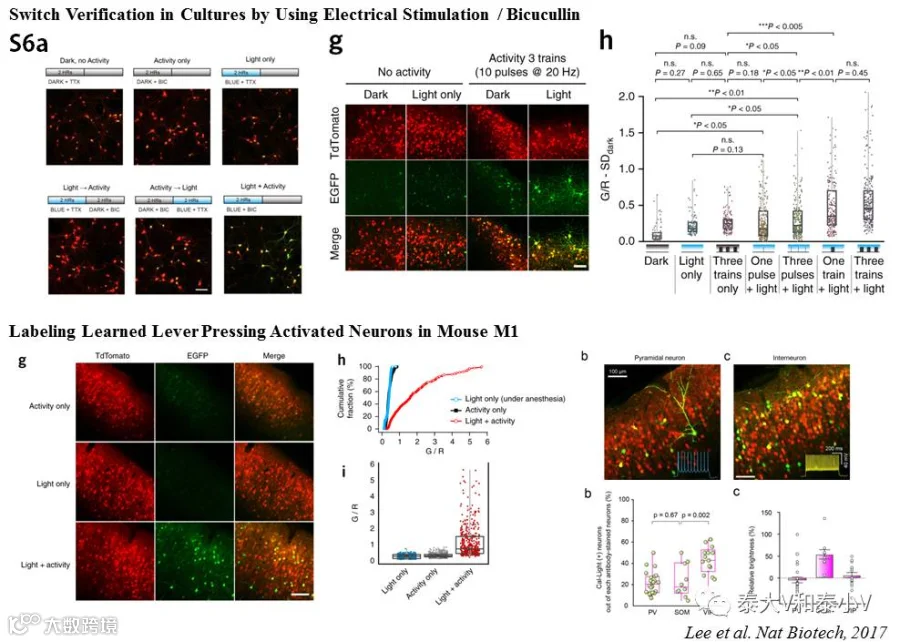
随后他们测试了Cal-Light在体标记行为相关神经集群的性能。先训练限水的小鼠通过连续压杆获得奖励性质的水,之后标记初级运动皮层M1在小鼠压杆训练(Lever-pressinglearning)时激活的神经集群。结果表明在体荧光标记表现出与离体测试一致的光及刺激共同依赖的特性(无光或是麻醉动物几无表达)。他们进一步让压杆学习所激活的M1神经集群表达嗜盐菌紫质eNpHR,随后利用光遗传实时沉默M1压杆学习相关的神经集群,即时地抑制了小鼠习得的压杆行为。而沉默小鼠走动(Walking)所激活的M1神经集群或是随机非同步沉默压杆学习激活的M1神经集群都不会影响小鼠的压杆行为,这表明Cal-Light的在体特异标记具有很高的时间分辨率(Lee et al. 2017)。
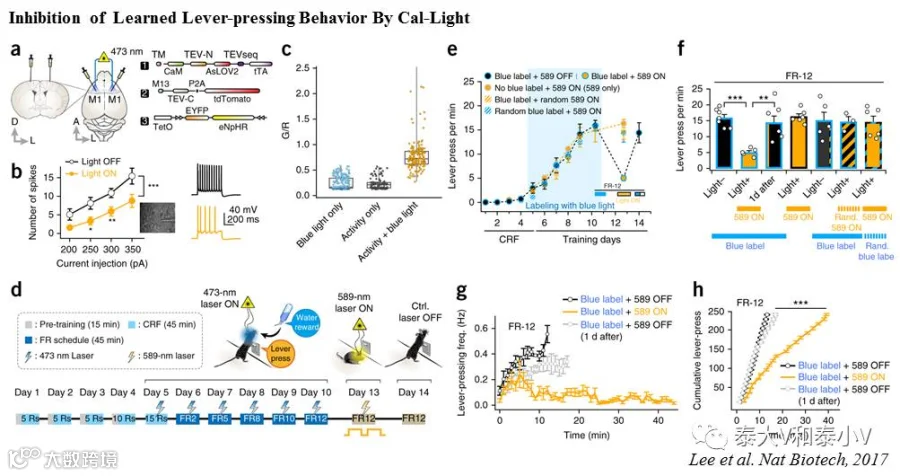
FLARE
与Cal-Light的工作机制类似,FLARE同样也是一个光控钙依赖组构式基因表达的系统。不同之处在于FLARE系统采取了一系列包括截短TEVp的C末端降低酶切亲和力太强引起的背景泄漏、通过易错PCR突变筛选得到光依赖特性强化版的eLOV及插入一段Nav1.6胞体靶向序列协助切下的tTA转位入核的方案来提高FLARE标记的信噪比。在大鼠培养神经元体外测试的研究表明,不论是电刺激还是更换高浓度Ca2+培养基引起的刺激都能实现差异显著的光控基因表达,随后他们结合钙成像及脑片电生理证实FLARE标记表达红移去极化视紫蛋白ChrimsonR的神经元可被红光再度激活。最后他们使用FLARE标记了小鼠跑转轮(Wheel running)时运动皮层被激活的神经集群,暂未提供FLARE在体功能验证的数据(Wang et al. 2017)。
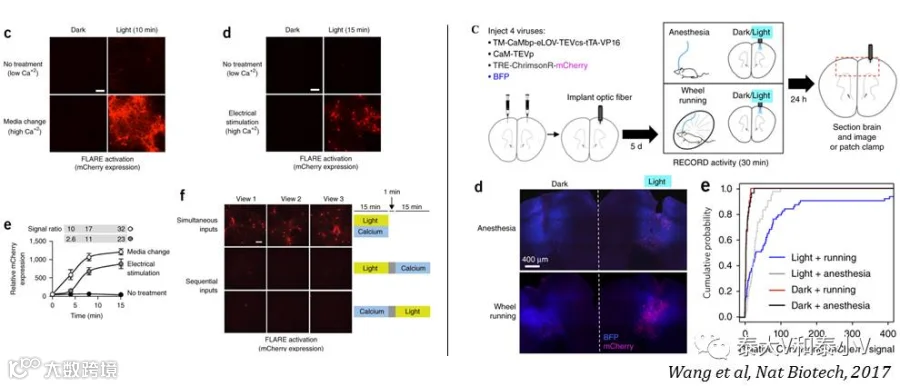
FLARE对多个元件进行的遗传改造显著提升了系统钙门控(Calcium gating)和光门控(Light gating)性能,离体测试表明FLARE光控信噪比高达120:1, 10min 高浓度与低浓度Ca2+刺激信号比能达到10:1。但他们也认识到在体实验中仍存在一些背景泄漏问题,以及通过病毒注射而后置入光纤给予蓝光进行诱导存在的不均质性,想必这也是Cal-Light系统一定程度上所共存的问题(见表9)。他们表示会进一步开发2.0版光控Ca2+依赖活性工具。
从1992年IEG研究的鼻祖罗氏分子生物学研究所的Tom Curran 和James I.Morgan开发第一个活动依赖的工具小鼠cfos-LacZ到最新的光控Ca2+依赖新工具Cal-Light/FLARE不过1/4个世纪的时间,但是关于记忆印迹及事件编码神经集群的研究日新月异。基于现存的活动依赖工具的特性,下面我们对工具应用进行简单概要:
多印迹/神经集群标记(Multi-stimuli Tagging)
大脑的功能编码通常是高度整合的,除了不同脑区及其环路主导不同的功能之外,也存在同一核团亚区被多种不同性质的刺激所激活从而编码多种信息的情形(Humphries et al. 2010,Lin et al. 2011, Janak & Tye. 2015)。同时标记多个印迹/神经集群为解析神经功能亚群(Subpopulations)提供了可能。自90年代末以来即有一系列基于IEG蛋白免疫组织化学染色(IHC)及mRNA荧光原位杂交(FISH)的双刺激标记方案的报道:
利用即早基因zif268激活产生的mRNA和蛋白质累积时程的差别,1997年Avi Chaudhuri实验室开发了I-FISH技术用以绘制视觉皮层响应两次间隔2h的视觉刺激的活性图谱(Chaudhuri et al. 1997,Zangenehpour et al. 2002):取脑时第一次刺激产生的蛋白处于高峰而mRNA已降解,第二次刺激产生的mRNA处于高峰但蛋白还未生成,因此两次刺激激活的神经集群可分别被IHC和FISH所表征。2014年Hailan Hu实验室开发了基于酪胺放大(TSA)系统的TAI-FISH技术进一步拓展了I-FISH双标技术的应用性,突破了NAc及LSv脑区存在的cfos IHC-FISH信号时程难以分离的限制,成功地绘制了边缘前脑情绪效价图谱(Xiu et al. 2014)。而John F. Guzowski实验室则于1999年和2002年分别开发了Arc catFISH(Guzowski et al. 1999)及Arc/homer1a catFISH(Vazdarjanova et al.2002)解析两次间隔20-30min的空间探索经历所激活的海马印迹。Arc catFISH利用生成的mRNA会随着时间由细胞核转位到细胞质的特点,通过64倍镜下信号细胞定位分析(Cellular compartment analysis)从而获得两次刺激的活性双标图谱;Arc/homer1a catFISH则是基于homer1a mRNA在刺激30min才开始生成而arc mRNA在5min左右即开始积累的特点分别对两次刺激进行标记。
通过活动依赖工具标记与IEG IHC相结合的方法进行双刺激标记最早被用于工具特异性检验(Liu et al. 2012, Kawashimaet al. 2013)和所标记的记忆印迹及神经集群功能编码特性的分析(Reijmers et al. 2007, Hanet al. 2007, Okuyama et al. 2016),随后亦被用于解析同一核团亚区存在不同的功能亚群(Gore et al. 2015, Sakuraiet al. 2016, Ye et al. 2016)以及记忆印迹信息编码的转移(Tayler et al. 2013, Kitamuraet al. 2017)。这一双标方案的优势在于无需进行操作复杂的原位杂交(ISH)实验(不过目前已有RNAscope技术可作为替代(Wang et al. 2012))且结果不受相继给予刺激可能存在干扰的影响。需要注意的是几乎所有IEG依赖的活性工具的外显率都低于50%,通过这一双标方案得到的结果可能是不全面的,另工具本身的忠实度及标记开放时间窗也极大程度决定了结果的可信度。
除了上述结合染色的方案,2016年Susumu Tonegawa实验室在研究腹侧海马CA1编码社交记忆的工作中将计时变色蛋白(Fluorescent timers)FT-Slow(Subach et al. 2009)引入了印迹双标系统(Okuyama et al. 2016)。利用红色荧光蛋白mCherry衍生的单分子突变体FT-Slow的发色团缓慢成熟(Chromophore formation)由蓝变红的特点,他们将AAV9-TRE-FT-Slow注入cfos-tTA小鼠的vCA1亚区,通过两次间隔72h的标记时间窗的开放实现了两次社交记忆印迹红色-蓝色荧光标记(小鼠A-A及小鼠A-B社交经历)。这种基于同一系统的标记方案极大地提升了标记的稳定性及可对比性。值得一提的是,FT-Slow也可应用于Cal-Light/FLARE系统进行双刺激标记检测工具的稳定性和特异性,以及多个神经亚群的表征。
多刺激标记扩展性能最强的是CANE,这一基于可降解受体的系统几乎可以进行无限次标记及载入,只需要每次刺激后注入带不同目的基因标签的包裹伪装病毒。CANE最大的突破在于不仅可以实现荧光标记表征,还可载入不同的效应蛋白如Cre, Flp, Dre等从而实现同一脑区多个神经亚群的分选及干预。当然,CANE, Cal-Light和FLARE都可以用于多个不同脑区的神经集群的捕获,只需要将包裹伪装病毒或是蓝光诱导(及病毒)同时/接连给予到不同的脑区即可,这对于从系统层面研究功能协调运作及信息流处理机制无疑是一项重大突破,为破译进而模拟复杂事件编码提供了一种可能。
印迹/神经集群环路解析(Circuit ArchitectureDeciphering)
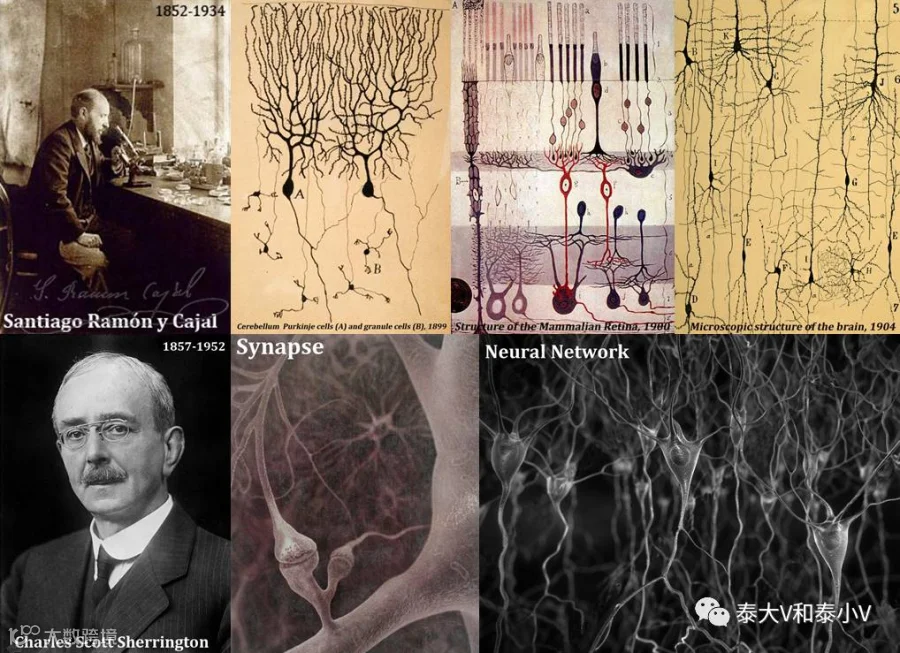
自19世纪末西班牙卓越的神经解剖学家卡哈尔(Santiago Ramón y Cajal)推行“神经元学说(Neuron doctrine)”及英国著名神经生理学家谢灵顿(Charles Scott Sherrington)提出“突触(Synapse)”这一阐释神经元连接单位的概念以来,神经环路及神经网络的研究异彩纷呈。
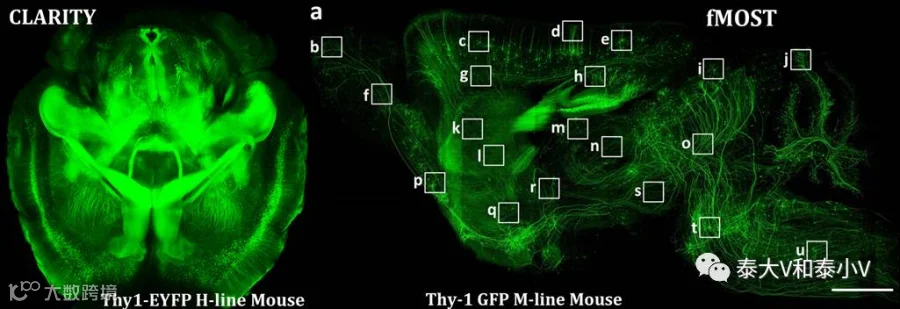
虽然在上世纪九十年代即已通过结合IEG IHC/ISH及下游注射逆行示踪剂(Fluorogold, Retrobeads, CTb)的方法来研究特定脑区事件相关神经集群的投射(Robertson et al. 1992,1995, Campeau et al. 2000),但真正实现相关环路的直接示踪还是来自于活动依赖工具的开发应用。从早期的tau-LacZ(Wilson et al. 2002, Reijmerset al. 2007)到近些年的荧光报告蛋白标记投射(Ye et al. 2016, Sakurai etal. 2016)为解析功能神经类群的环路基础开辟了道路。而通过膜结合版GFP(mGFP)或是EYFP-NRN显示整个神经纤维以及利用融合了荧光蛋白的小突触泡蛋白(Synaptobrevin)或是突触小泡蛋白(Synaptophysin)指示突触前所在位置的方案也可应用于神经集群的环路示踪,实现更高效而精准的投射及突触的标记(SynaptoTag)(Xu & Südhof. 2013, Ohet al. 2014, Lerner et al. 2015, Ye et al. 2016)。此外近些年飞速发展的全脑3D成像技术如基于组织透明技术及光切片的BBAB, Scale, 3DISCO, ClearT,CLARITY,SeeDB, CUBIC, iDISCO, PACT/PARS, SWITCH等(Lee et al. 2016)、荧光显微光学切片断层成像技术(fMOST)(Li et al. 2010, Gong et al.2016)、连续双光子组织断层扫描(STP)(Ragan et al. 2012, Kim etal. 2015)及高速断面连续断层扫描技术(FAST)(Seirik et al. 2017)则为重构神经集群精细的三维结构(Circuit architecture)提供了可能。另CLARITY及iDISCO等组织透明处理后的脑子还可进行IHC及ISH进一步检测亚群存在的分子特性(Chung et al. 2013, Yeet al. 2016, Renier et al. 2016)。
除了解析投射输出,利用伪装改造的单突触(Monosynaptic)RV(Wickersham et al. 2007)进行逆向跨突触追踪活动相关神经集群的输入同样也有所报道(Sakurai et al. 2016, Kitamuraet al. 2017)。随着减毒高效版RV病毒株CVS-N2cDG(Reardon et al. 2016)及自失活版RV-SiR(Ciabatti et al. 2017)的开发应用,单突触输入的操纵及监测也成为了可能,这为研究功能神经网络及其信息流开辟了道路。
印迹/神经集群分子基础检测(Molecular Features Uncovering)
神经集群特殊的环路结构为功能编码及信息流处理提供了运作支架,然而印迹/神经集群特定功能的编码必然依赖于其特定的分子基础。此外相对于正常生理情况,病理状态如成瘾、神经精神疾病等通常也表现为特定神经类群的分子改变(Molecular alteration)(Cruz et al. 2013)。破解神经集群所存在的分子基础对于功能解析及开发药物干预靶点无疑具有重大意义。检测活性神经类群的分子特性及变化最早的工作是Bruce T. Hope实验室基于荧光激活细胞分选术(FACS)的一系列研究报道(见表10)。
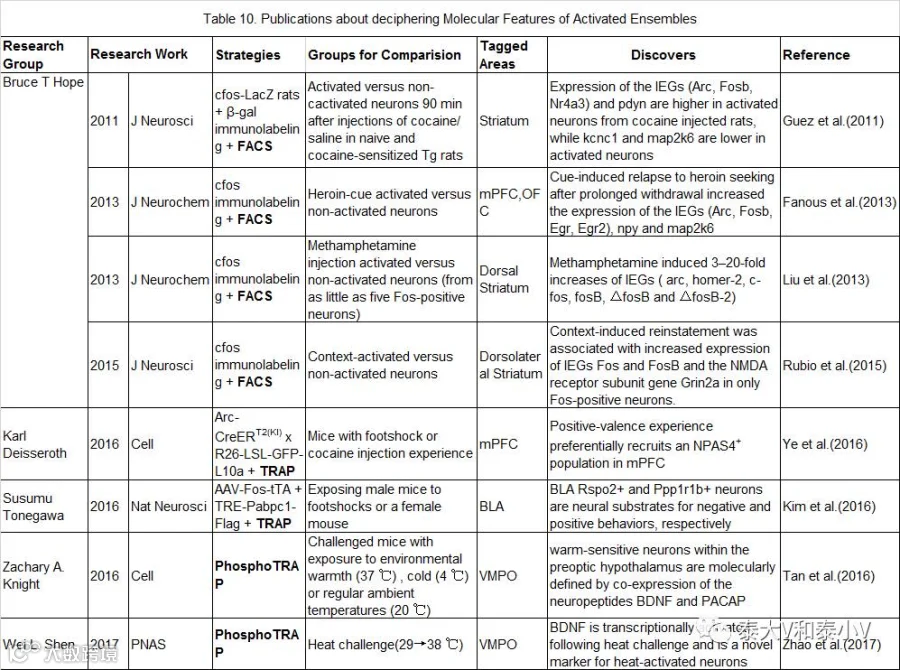
他们通过免疫荧光标记(Immunolabeling)纹状体/前皮层被成瘾药物或是偶联的线索/环境所激活的细胞(cfos-LacZ大鼠β-gal+或是SD大鼠cfos+)从而进行FACS分选,之后利用微阵列(Microarray)及定量PCR(qPCR)检测基因的转录水平。由于检测的文库太小(10-20个基因),得到的结果相对单一,但这都是一些有益的探索(Guez et al. 2011, Fanous etal. 2013, Liu et al. 2013, Rubio et al. 2015)。
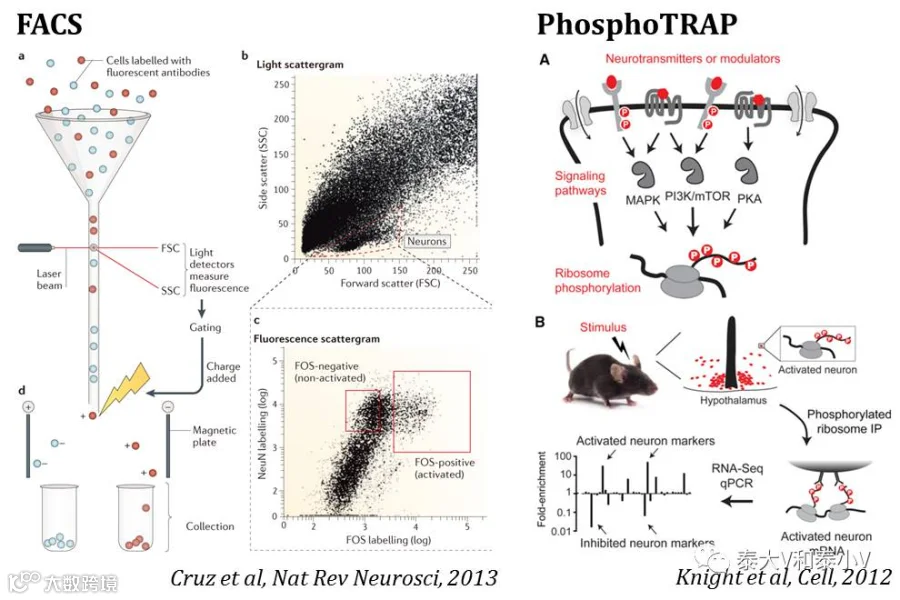
除了上述的基于流式细胞术(Flow cytometry)的分选方案,近些年更广为采用的是基于免疫沉淀反应(Immunoprecipitation, IP)的翻译核糖体亲和纯化技术(Translating Ribosome AffinityPurification, TRAP)。利用活动依赖工具如Arc-CreERT2(KI)x R26-LSL-GFP-L10a杂交小鼠使其活性神经集群的核糖体L10a亚基融合表达GFP(Ye et al. 2016)或是注射AAV9-Fos-tTA及AAV9-TRE-Pabpc1-Flag病毒的小鼠活化细胞表达抗原表位带Flag标签的RNA poly(A)结合蛋白(可进一步结合于转录生成的mRNA)(Kim et al. 2016),随后利用GFP/Flag抗体将携带了翻译中的mRNA的核糖体/结合蛋白免疫沉淀下来,之后进行RNA测序分析。两项工作分别筛选了mPFC和BLA正/负面刺激响应神经集群所存在的分子标记。而加州大学洛杉矶分校(UCSF)的Zachary A. Knight在其博后期间所开发的PhosphoTRAP技术(Knight et al. 2012)则是利用活化细胞核糖体S6亚基本身存在磷酸化的特点,利用高效而忠实的pS6抗体(抗体是TRAP成败的关键)直接进行TRAP即可。Zachary A. Knight实验室及Wei L. Shen实验室通过PhosphoTRAP成功地筛出了腹内侧视前区前部(VMPO)热感应神经元存在的分子标记(Tan et al. 2016, Zhaoet al. 2017)。此外单细胞测序(Single-cell RNA-seq)技术结合活性工具标记也可用于印迹/神经集群的分子基础检测(Molecular profiling)。值得注意的是以上工作仅表明所筛出的分子可作为功能类群的标签,但尚未见到有干预筛选所得的分子直接影响功能的报道。
展望
应用活动依赖工具目前已有一系列关于记忆印迹、感觉及情绪效价、行为及行为异常编码的研究工作报道,而新工具更是可以用于研究精细行为及复杂功能的编码。但开发更优的面向未来的活动依赖工具的工作将永无止境,探索其他条件性基因表达策略与之前的工具联用或是择优替换不无可能。例如Frank Edenhofer实验室开发的Mifepristone所激活的Cre*PR拥有比CreERT2更低的背景泄漏(Wunderlich et al. 2001),而Anton Maximov实验室则将Cre与降解结构域(DD)融合成DD-Cre,使得只有在trimethoprim存在的情况下才能阻止降解从而实现快速诱导(Sando et al. 2013)。
另需要明确的是利用激活所表征的神经集群只是功能编码图式的一部分,而抑制同样在里面扮演了重要角色。虽然Ca2+指示剂在一定程度上能够表征神经元的抑制现象,但开发可直接表征神经元兴奋抑制状态的电压敏感指示剂(Marshall et al. 2016)等的工作一直令业界翘首。只要能够开发出合用的指示剂,被抑制的神经集群也同样可以被捕获。
细胞水平的在体成像技术监测神经集群活动与高分辨率活动依赖工具操纵神经集群活性相结合是未来的一个重要方向。但不得不承认我们现在所能实现的活性模拟及功能复现是相对的,机体进行特定的生理功能输出时几乎不太可能像特定的光遗传给光程序一齐调制整个神经集群那样地工作。虽然现已开发成像与记录相结合的光遗传闭环给光模拟生理激活(Closed-loop optogenetics)的方案(Akerboom et al. 2013, Grosenicket al. 2015),甚至于开发了基于3D钙成像(Volumetric imaging)的双光子全息光遗传(Holographic optogenetics)系统解析神经集群(dal et al. 2017, Carrilloet al. 2017),但距离破解认知(Cognition)与意识(Consciousness)、智力(Intelligence)与情感(Emotion)、记忆(Memory)与社会行为(Social behavior)等一系列永恒命题的生物学机制,还有漫长的路要走。一切生命的奥秘所在,即是众妙之门,等待着一代又一代的人去开启和穿越。
致谢
感谢生命里的缪斯H.H & ZHE; 感谢SHEN (ZJU), JIE(ZJU), GANG (ZJU), NI (FU), Yi (ION), Dafeng (Taitool), Kunming(Newdoon)及Cihang (Newdoon)对文稿所提供的修改意见。
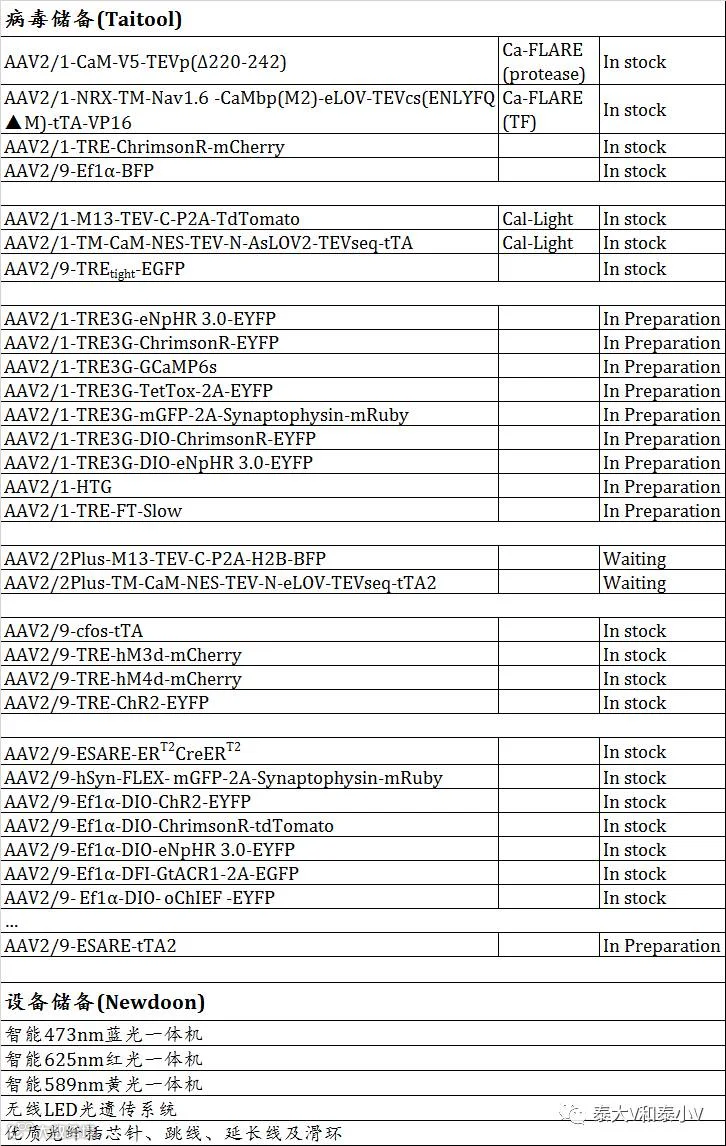
兵库蓄势,粮草先行。上海泰儿图生物科技有限公司(Taitool)与杭州纽顿科技有限公司(Newdoon)联合启动“Ac Action!”计划(详见相应推文),诚邀您华山论剑,赏味电光火石,共修活性工具兵器谱!


参考文献
