


一、背景简介:为什么我们需要“看见”特定的脑细胞?


二、从分子身份证到细胞开关:数据驱动的特异性靶向工具开发策略 |

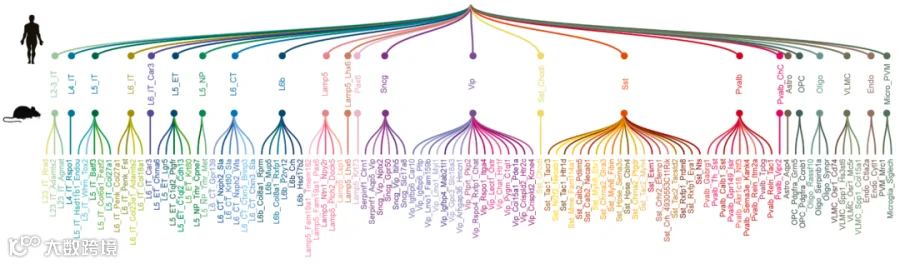
图1 小鼠-人类皮质细胞亚类分类法的简图,
以及仅针对小鼠的聚类级别分类法



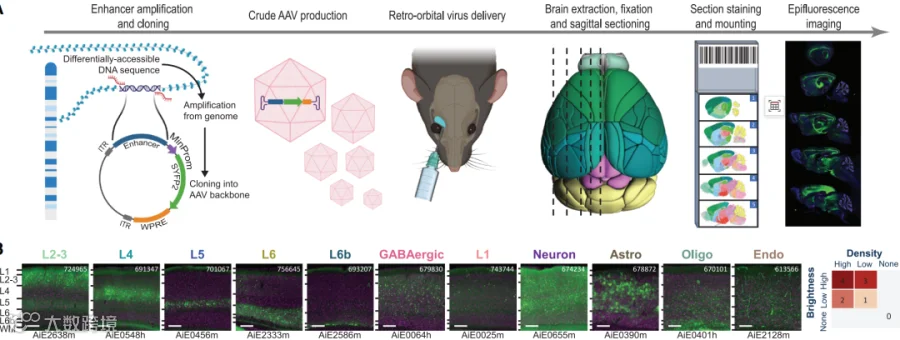
图2 增强子筛选流程和
视觉皮层的代表性图像

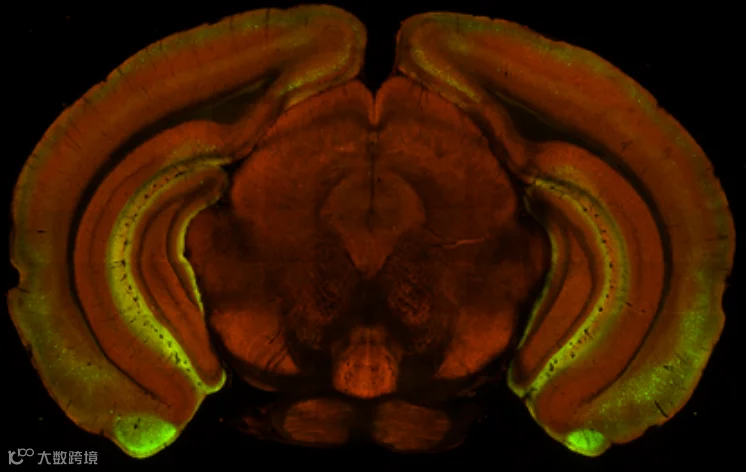
图3 AiE0680m-SYFP2-RO的STPT图像
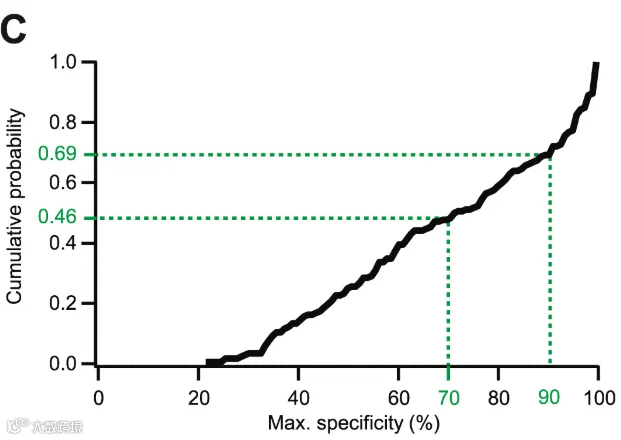
图4 增强子累积分布图

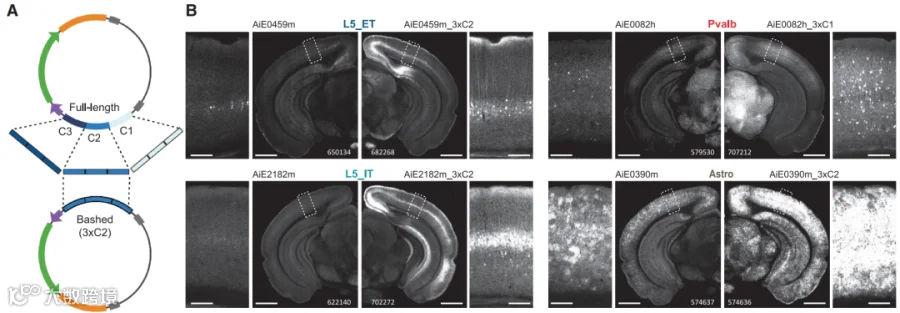
图5 增强子核心优化示意图及实验结果
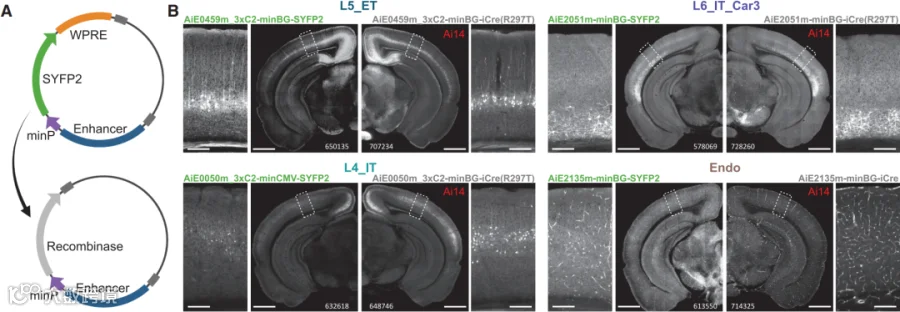
图6 重组酶设计示意图及实验结果
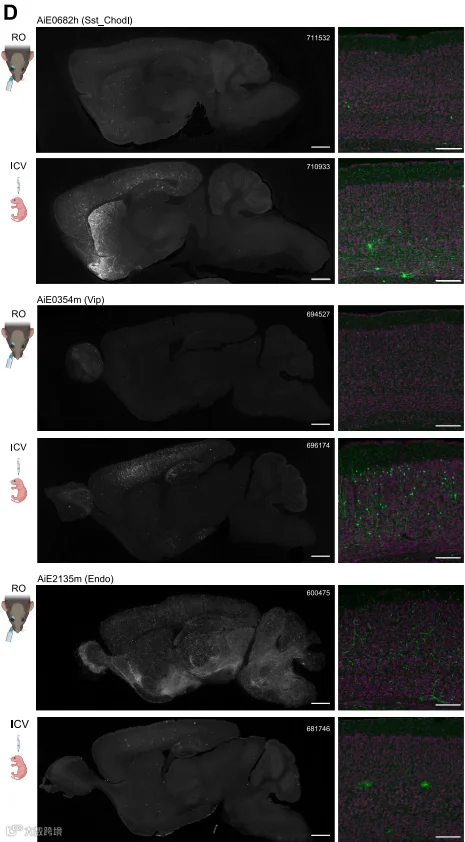
图7 三种增强子AAVs不同递送方式
(RO和ICV)表达差异
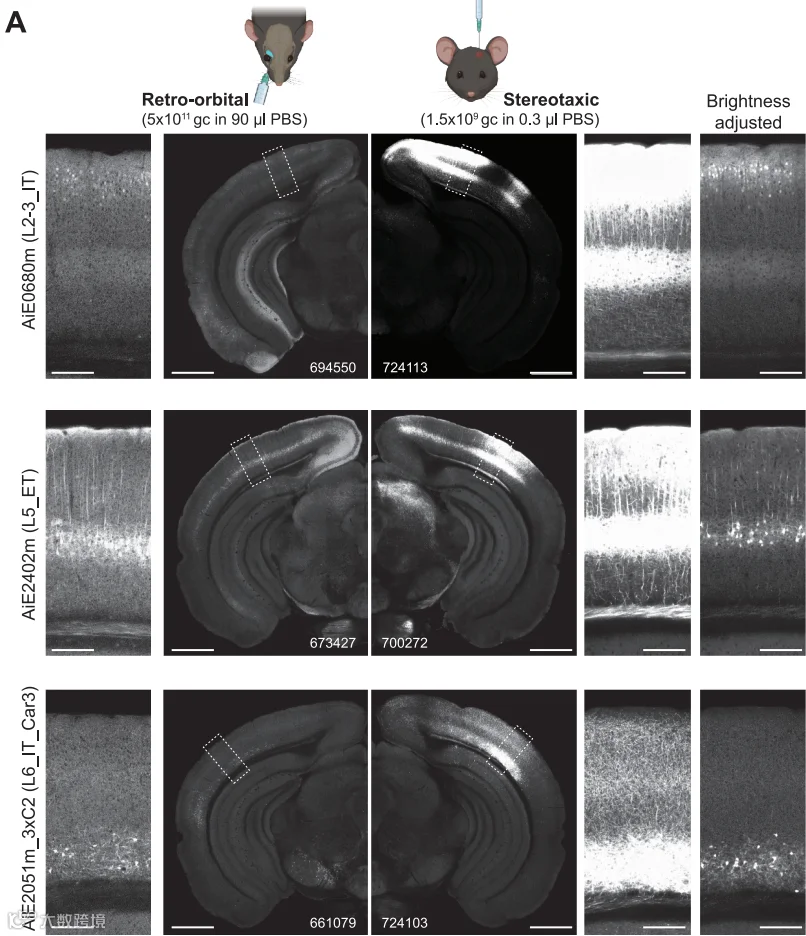
图8 三种增强子AAVs不同递送方式
(RO和STX)表达差异
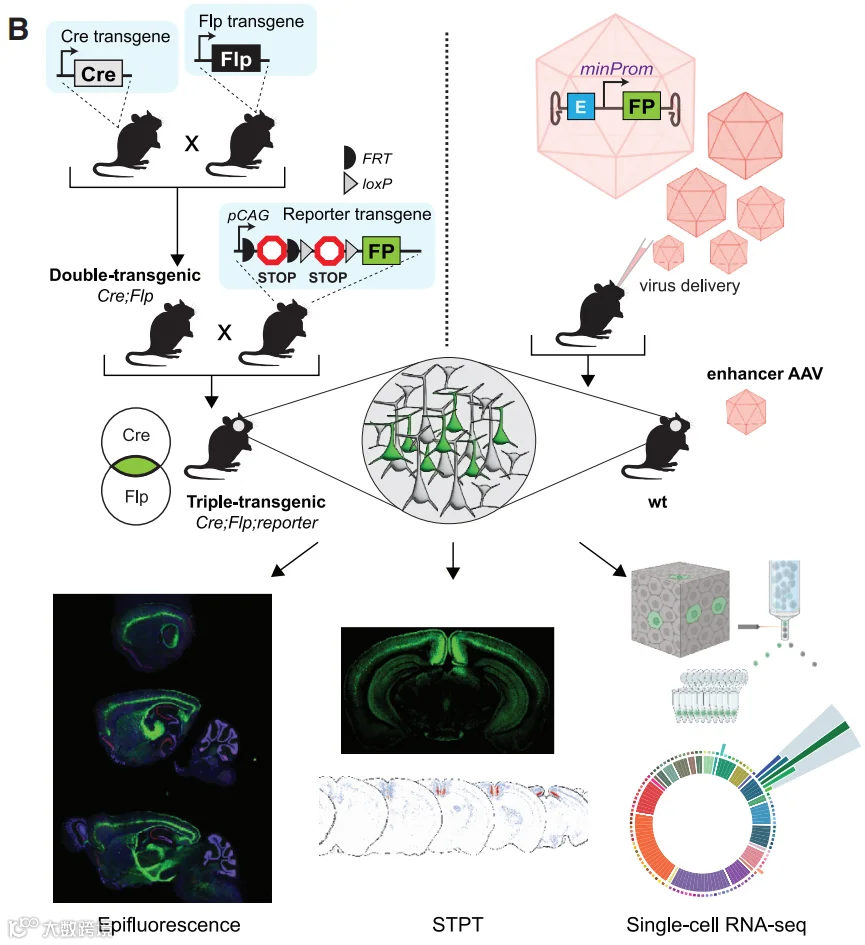
图9 基因编辑鼠的生成和表征

三、实际应用:这套工具将如何改变神经科学研究?
神经环路解析:研究人员可以精确标记特定类型神经元,用荧光追踪其长长的轴突投射,绘制出前所未有的精细“大脑接线图”。结合光遗传学或化学遗传学,可以激活或抑制这些细胞,实时观察其对动物行为、脑电活动的影响,从而直接验证其功能。

四、最后总结:开启大脑细胞类型研究的新纪元

艾伦实验室的这项工作,不仅仅是为学术界贡献了上千个新工具,更重要的是它展示并实践了一种规模化、系统化、数据驱动的现代神经科学研究范式。
过去,寻找一个可用的细胞类型特异性工具往往依赖运气和零散报道。现在,研究者可以基于统一的细胞分类学图谱,在一个集中、经过严格验证的资源库中,理性地选择和组合工具。这种从“手工作坊”到“标准化大生产”的转变,将极大地提升研究效率、可重复性和结果的可靠性。
当然,研究也指出了应用中的注意事项,如病毒滴度、递送方法对结果的影响,以及不同细胞类型对病毒转导敏感性的差异等。这些提醒研究者,即使是“名人堂”工具,也需要在自己的实验体系中,用多种模态(如成像和测序)进行验证。

根据“名人堂”工具以及实验结果,我们提供一系列现货工具:
增强子ID
|
元件
|
注射方式
|
靶细胞类型
|
AiE0680m
|
pAAV-ePro(AiE0680m)-EGFP-WPRE-pA
|
RO
|
L2-3_IT
|
AiE0680m
|
pAAV-ePro(AiE0680m)-iCre(R297T)-WPRE-pA
|
RO
|
L2-3_IT
|
AiE2543m
|
pAAV-ePro(AiE2543m)-iCre(R297T)-WPRE-pA
|
RO
|
L2-3_IT
|
AiE2543m
|
pAAV-ePro(AiE2543m)-EGFP-WPRE-pA
|
RO,STX
|
L2-3_IT
|
AiE0050m_3xC2
|
pAAV-ePro(AiE0050m_3xC2)-iCre(R297T)-WPRE-pA
|
RO
|
L4_IT
|
AiE0050m_3xC2
|
pAAV-ePro(AiE0050m_3xC2)-EGFP-WPRE-pA
|
RO
|
L4_IT
|
AiE0671m_3xC2
|
pAAV-ePro(AiE0671m_3xC2)-iCre(R297T)-WPRE-pA
|
RO
|
L4_IT
|
AiE0671m_3xC2
|
pAAV-ePro(AiE0671m_3xC2)-EGFP-WPRE-pA
|
RO,STX
|
L4_IT
|
AiE2182m_3xC2
|
pAAV-ePro(AiE2182m_3xC2)-iCre(R297T)-WPRE-pA
|
RO
|
L5_IT
|
AiE2182m_3xC2
|
pAAV-ePro(AiE2182m_3xC2)-EGFP-WPRE-pA
|
RO,STX
|
L5_IT
|
AiE0456m
|
pAAV-ePro(AiE0456m)-iCre(R297T)-WPRE-pA
|
RO
|
L5_ET
|
AiE0456m
|
pAAV-ePro(AiE0456m)-EGFP-WPRE-pA
|
RO
|
L5_ET
|
AiE0459m_3xC2
|
pAAV-ePro(AiE0459m_3xC2)-iCre(R297T)-WPRE-pA
|
RO
|
L5_ET
|
AiE0459m_3xC2
|
pAAV-ePro(AiE0459m_3xC2)-EGFP-WPRE-pA
|
RO
|
L5_ET
|
AiE0464m
|
pAAV-ePro(AiE0464m)-EGFP-WPRE-pA
|
RO,STX
|
L5_ET
|
AiE0674m
|
pAAV-ePro(AiE0674m)-EGFP-WPRE-pA
|
RO
|
L5_ET
|
AiE2402m
|
pAAV-ePro(AiE2402m)-EGFP-WPRE-pA
|
RO,ICV
|
L5_ET
|
AiE0667m
|
pAAV-ePro(AiE0667m)-EGFP-WPRE-pA
|
RO
|
L5_ET_Chrna6
|
AiE2449m
|
pAAV-ePro(AiE2449m)-EGFP-WPRE-pA
|
RO
|
L5_ET_Chrna6
|
AiE2051m
|
pAAV-ePro(AiE2051m)-iCre(R297T)-WPRE-pA
|
RO
|
L6_IT_Car3
|
AiE2051m
|
pAAV-ePro(AiE2051m)-EGFP-WPRE-pA
|
RO
|
L6_IT_Car3
|
AiE0772m
|
pAAV-ePro(AiE0772m)-EGFP-WPRE-pA
|
RO
|
L6_IT_Car3
|
AiE2586m
|
pAAV-ePro(AiE2586m)-iCre(R297T)-WPRE-pA
|
RO
|
L6b
|
AiE2586m
|
pAAV-ePro(AiE2586m)-EGFP-WPRE-pA
|
RO,STX
|
L6b
|
AiE2587m
|
pAAV-ePro(AiE2586m)-iCre(R297T)-WPRE-pA
|
RO
|
L6b
|
AiE2587m
|
pAAV-ePro(AiE2587m)-EGFP-WPRE-pA
|
RO
|
L6b
|
AiE0724m
|
pAAV-ePro(AiE0724m)-iCre(R297T)-WPRE-pA
|
RO
|
Lamp5
|
AiE0724m
|
pAAV-ePro(AiE0724m)-EGFP-WPRE-pA
|
RO
|
Lamp5
|
AiE2103m
|
pAAV-ePro(AiE2103m)-iCre(R297T)-WPRE-pA
|
RO
|
Lamp5
|
AiE2103m
|
pAAV-ePro(AiE2103m)-EGFP-WPRE-pA
|
RO
|
Lamp5
|
hsA2-AiE0219h
|
pAAV-ePro(hsA2-AiE0219h)-iCre(R297T)-WPRE-pA
|
RO
|
Vip
|
hsA2-AiE0219h
|
pAAV-ePro(hsA2-AiE0219h)-EGFP-WPRE-pA
|
ICV
|
Vip
|
3xSP10ins-AiE0354h
|
pAAV-ePro(3xSP10ins-AiE0354h)-iCre-WPRE-pA
|
RO
|
Vip
|
3xSP10ins-AiE0354h
|
pAAV-ePro(3xSP10ins-AiE0354h)-EGFP-WPRE-pA
|
RO,ICV
|
Vip
|
3xSP10ins-AiE0354m
|
pAAV-ePro(3xSP10ins-AiE0354m)-EGFP-WPRE-pA
|
ICV
|
Vip
|
AiE0682h
|
pAAV-ePro(AiE0682h)-iCre(R297T)-WPRE-pA
|
RO
|
Sst_Chodl
|
AiE0682h
|
pAAV-ePro(AiE0682h)-EGFP-WPRE-pA
|
RO
|
Sst_Chodl
|
AiE0600m
|
pAAV-ePro(AiE0600m)-EGFP-WPRE-pA
|
RO
|
Sst_Chodl
|
AiE0519h
|
pAAV-ePro(AiE0519h)-EGFP-WPRE-pA
|
RO,ICV
|
Sst
|
AiE0170h_3xC2
|
pAAV-ePro(AiE0170h_3xC2)-EGFP-WPRE-pA
|
RO
|
Sst
|
hsA2-AiE0140h
|
pAAV-ePro(hsA2-AiE0140h)-iCre(R297T)-WPRE-pA
|
RO
|
Pvalb
|
hsA2-AiE0140h
|
pAAV-ePro(hsA2-AiE0140h)-EGFP-WPRE-pA
|
RO
|
Pvalb
|
hsA2-AiE0086m
|
pAAV-ePro(hsA2-AiE0086m)-EGFP-WPRE-pA
|
RO
|
Pvalb
|
AiE0475m
|
pAAV-ePro(AiE0475m)-EGFP-WPRE-pA
|
RO,ICV
|
Pvalb_ChC
|
AiE0390m_3xC2
|
pAAV-ePro(AiE0390m_3xC2)-iCre(R297T)-WPRE-pA
|
RO
|
Astro
|
AiE0390m_3xC2
|
pAAV-ePro(AiE0390m_3xC2)-EGFP-WPRE-pA
|
RO
|
Astro
|
AiE0390h_3xC2
|
pAAV-ePro(AiE0390h_3xC2)-EGFP-WPRE-pA
|
RO
|
Astro
|
AiE0387m
|
pAAV-ePro(AiE0387m)-iCre(R297T)-WPRE-pA
|
RO
|
Astro
|
AiE0387m
|
pAAV-ePro(AiE0387m)-EGFP-WPRE-pA
|
RO,ICV
|
Astro
|
AiE0395h
|
pAAV-ePro(AiE0395h)-EGFP-WPRE-pA
|
RO
|
Oligo
|
AiE0397m
|
pAAV-ePro(AiE0397m)-EGFP-WPRE-pA
|
RO
|
Oligo
|
AiE2135m
|
pAAV-ePro(AiE2135m)-iCre-WPRE-pA
|
RO
|
Endo
|
AiE2135m
|
pAAV-ePro(AiE2135m)-EGFP-WPRE-pA
|
RO,ICV
|
Endo
|
AiE2128m
|
pAAV-ePro(AiE2128m)-EGFP-WPRE-pA
|
RO
|
Endo
|
注:靶细胞类型分类说明:
基本特点:释放谷氨酸(Glutamate)作为神经递质,主要起兴奋性作用,构成皮层神经网络的主要连接基础。
常见亚型(按皮层分层):
L2-3_IT(Layer 2/3 Intratelencephalic):
位置:主要位于皮层第2/3层。
投射:投射至同侧和对侧皮层(通过胼胝体或前联合),也投射至纹状体。
形态:薄顶树突,轴突主要下降至L5。
电生理:静息膜电位较超极化,发放稀疏。
功能:参与皮层间信息整合与传递,尤其是感觉和运动信息的初步处理。
L4_IT:
位置:主要位于皮层第4层(感觉皮层)。
投射:投射至L2/3 IT神经元、同层的其他L4神经元,少量投射到皮层下的纹状体。
形态:包括锥体神经元、星状锥体神经元、棘星状神经元等亚型。
功能:是丘脑输入的主要接收层,参与初级感觉信息处理。
备注:运动皮层(MOp)无明显的L4细胞构筑,但存在类似L4的神经元。
L5_IT:
位置:主要位于皮层第5层。
投射:比L2-3 IT更广泛,包括对纹状体的更强投射。
电生理:比L2-3 IT更活跃,动作电位发放更频繁。
功能:参与皮层-纹状体通路,调控运动与行为规划。
L5_ET(Layer5 extratelencephalic projecting):
别名:锥体束神经元(Pyramidal Tract, PT)或皮层下投射神经元。
投射:投射至皮层下结构,如丘脑、中脑、脑干、脊髓。
形态:厚顶树突,轴突长距离投射。
电生理:静息膜电位较去极化,输入电阻低,动作电位易爆发。
功能:参与运动输出、觉醒调控、自主神经功能。
L6_IT_Car3:
位置:皮层第6层,表达Carbonic Anhydrase 3(Car3)。
投射:广泛的皮层内投射,但无纹状体侧支投射。
功能:参与局部深层神经元之间的信息交换。
L6b:
位置:第6层底部神经元,位于皮层最深层。
投射:主要投射至局部和邻近皮层的L1。
基本特点:释放GABA作为神经递质,抑制其他神经元活动,维持神经网络平衡,防止过度兴奋。
常见亚型:
Lamp5:
标记基因:Lamp5、Pdlim5。
位置:各层均有,L1层尤为突出。
形态:多极树突,高密度的胞周轴突丛。
电生理:多为晚期发放型,使用体积传递(volume transmission)。
功能:通过GABA_B受体介导慢抑制,参与抑制性调控与神经调节。
Vip(Vasoactive Intestinal Polypeptide)神经元:
投射:主要靶向其他GABA能神经元(尤其是SST神经元),从而“去抑制”锥体神经元。
形态:双极/双簇状树突,轴突下降至L6/白质边界。
功能:接收高阶皮层和皮层下神经调控输入,参与注意力、觉醒、行为状态调节。
Sst_Chodl:
亚型:属于Sst类,可能为特定分子标记亚型(如Chodl+)。
功能:推测具有特化的抑制性调控功能,具体功能仍需进一步研究。
Sst(Somatostatin)神经元:
亚型:包括Martinotti细胞和非Martinotti细胞。
投射:Martinotti细胞投射至L1,非Martinotti细胞多投射至L4。
功能:参与反馈抑制、调控锥体神经元远端树突整合。
Pvalb(Parvalbumin)神经元:
亚型:主要包括篮状细胞和Chandelier细胞。
电生理:快速发放型(fast-spiking)。
功能:调控皮层兴奋-抑制平衡、参与γ振荡、增益控制。
Pvalb_ChC(Chandelier Cell):
形态:轴突呈烛台状,靶向锥体神经元轴突起始段。
基本特点:不参与电信号传导,但在神经网络中起支持、保护、代谢调控等重要作用。
常见类型:
Astro(Astrocytes):星形胶质细胞,参与血脑屏障维护、突触传递调控、神经元代谢支持等。
Oligo(Oligodendrocytes):少突胶质细胞,形成髓鞘包裹轴突,加快神经冲动传导速度。
[1]Ben-Simon Y, et al. A suite of enhancer AAVs and transgenic mouse lines for genetic access to cortical cell types. bioRxiv [Preprint]. 2024 Sep 26:2024.06.10.597244.
[2]Mao X, Staiger JF. Multimodal cortical neuronal cell type classification. Pflugers Arch. 2024 May;476(5):721-733.
END


扫描二维码
获取更多信息

点击蓝字关注我们吧!!