


大家好呀,自从前段时间泰儿图的网红新品 —— mDlx 增强子
上市以后,收到了来自实验室前方的最新回馈数据,为标记GABA能中间神经元而生的新晋网红究竟好不好用?一起来看看!
话不多说,先看图
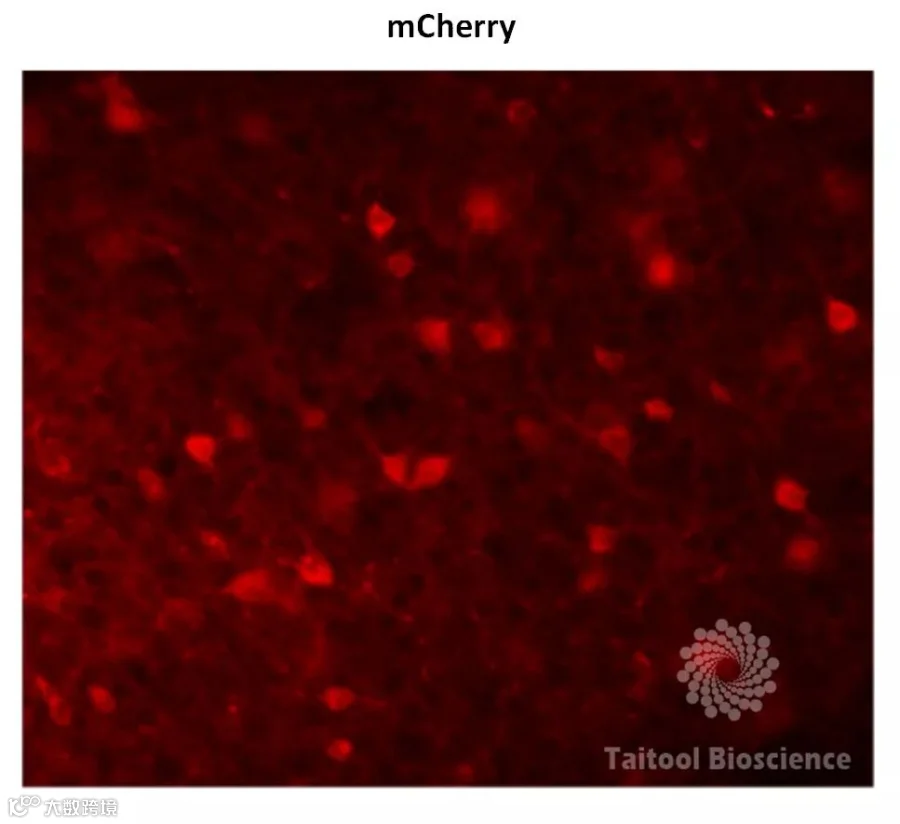
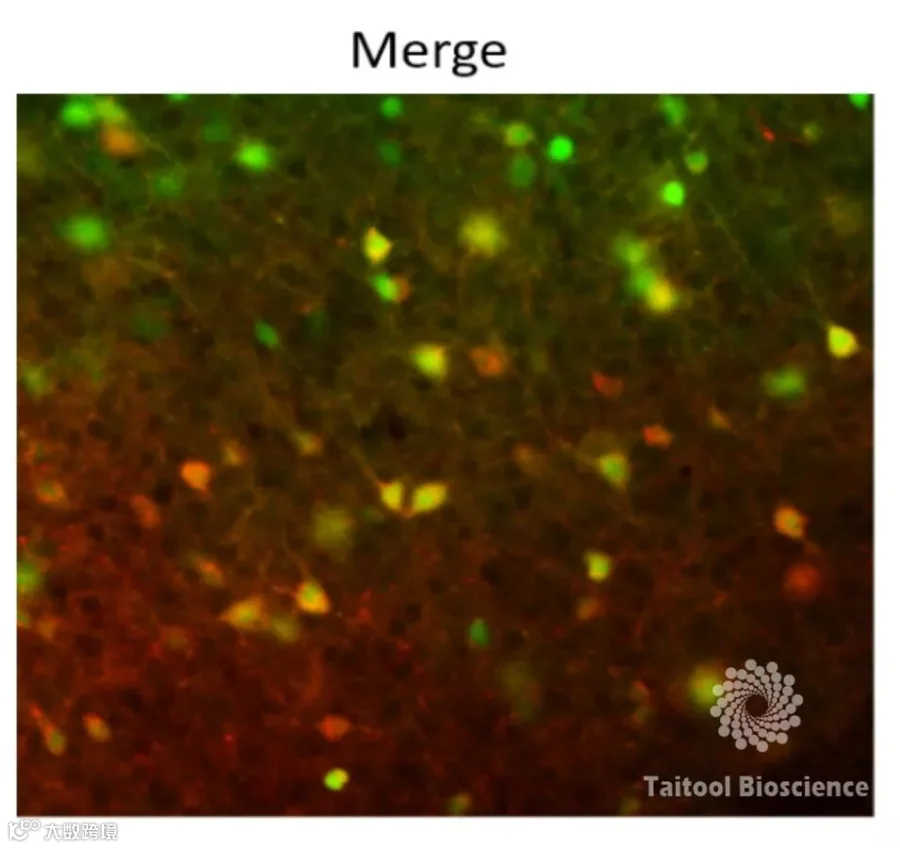


实 验 信 息
病毒载体:AAV2/9-mDlx-oChIEF-mCherry-ER2;
泰儿图货号:S0511-9
血清型:AAV2/9
注射区域:皮层
注射体积:500 nL
载体滴度: 2.8E+12 V.G./mL
表达时间:16天
实验结果:红色为病毒表达的mCherry,绿色为GAD67信号,结果显示mCherry的信号几乎100%与绿色共标,说明该载体特异性地感染了GABA能神经元。
该结果与上期推文中J.Dimidschstein等人的实验验证的类似,携带mDlx增强子的新型AAV病毒载体,它能限制基因表达到GABA能中间神经元。这种载体的表达强效又具有特异性,可以实现形态学可视化,时事动态检测和操纵小鼠等多种物种的中间神经元。
mDlx现今科研地位如何?
如果说到了现在,那就不能不讲讲以往常用的识别和操纵GABA能神经元策略了:
1. 利用cre重组酶驱动报告基因的方法借助AAV病毒标记GABA能细胞。但是单一的基因和单一的细胞类型可能并不是特别匹配,多数情况下是单一基因下标记多个细胞类群。
2. 利用cre,Flp或其他重组酶的交叉策略,提高细胞定位的精确度,共标及非共标的细胞类群;但是现有的多重组合还是很有限的。
3. 转录因子通常只在祖细胞中起作用,在成熟的神经元中会减少表达甚至改变表达模式。
4. 利用现有的能定位GABA能神经元的转基因小鼠,做这些细胞的基因的表达谱,有可能定位其中更少的细胞类型。
mDlx增强子推出的意义也就变得明确:
1. 突破了在非转基因动物中不能精确识别和操纵GABA能神经元类型的限制。
2. AAV装载容量很有限(~4.7kb),只能通过短小的顺势调控序列限制在特定的细胞中表达。而增强子是包含转录因子结合位点的顺式作用元件,其负责基因表达的组织特异性转录调控。
3. Dlx作为一个有效的工具,被有效证明可用来研究多种物种的海马,皮层和纹状体的抑制性GABA功能,甚至在临床研究中都具有有较强的潜在效用。

本次AAV2/9-mDlx-oChIEF-mCherry-ER2载体,mDlx增强子结合了oChIEF的首秀表现相当出色,但是话说oChIEF是个什么东西呢?
oChIEF的前身是ChIEF,是ChR1和ChR2两个蛋白的融合体,两者同时在170位的Ile突变为Val。这个蛋白融合体的特点是在长时间光照下,平台期电流和峰值电流的比值较大,那么也就意味着光照下通道的失活较少,适合长时间刺激。而Ile的突变加快了通道的关闭速度,使得ChIEF适合做高频的刺激使用。Karl实验室比较过用ChIEF和hChR2(E123A)在parvalbumin在神经元中做高频刺激,最终证明两者的效果相当。而oChIEF,就是在ChIEF基础上经过密码子优化的版本。听上去是不是有点像牛A + 牛C之后变成了他们中间的那个家伙呢?

AAV2/9-mDlx-oChIEF-mCherry-ER2
同款AAV载体现在你也可以同样可以拥有!
不仅oChIEF!
还有hM3D(Gq)! ChrimsonR! NpHR! hGtACR1! hChR2(H134R)! ...
更多实力目的基因搭配等你体验!
满1500起,还送快乐水!今夏就要你快乐!

话说聪明的少年已经去喝着快乐水美滋滋看实验数据了~
参考文献
[1] Dimidschstein J, Chen Q, Tremblay R, et al. A viral strategy for targeting and manipulating interneurons across vertebrate secies[J]. Nature neuroscience, 2016, 19(12): 1743.
[2] Taniguchi, H., He, M., Wu, P., Kim, S., Paik, R., Sugino, K., Kvitsiani, D., Fu, Y., Lu, J., Lin, Y., et al. (2011). A resource of Cre driver lines for genetic targeting of GABAergic neurons in cerebral cortex. Neuron 71, 995–1013.
[3] Dymecki, S.M., and Kim, J.C. (2007). Molecular neuroanatomy’s ‘‘Three Gs’’: a primer. Neuron 54, 17–34.
[4] Dymecki, S.M., Ray, R.S., and Kim, J.C. (2010). Mapping cell fate and function using recombinase-based intersectional strategies. Methods Enzymol. 477, 183–213.
[5] Fenno L E, Mattis J, Ramakrishnan C, et al. Targeting cells with single vectors using multiple-feature Boolean logic[J]. Nature methods, 2014, 11(7): 763.
[6] Madisen L, Garner A R, Shimaoka D, et al. Transgenic mice for intersectional targeting of neural sensors and effectors with high specificity and performance[J]. Neuron, 2015, 85(5): 942-958.
[7] He M, Tucciarone J, Lee S H, et al. Strategies and tools for combinatorial targeting of GABAergic neurons in mouse cerebral cortex[J]. Neuron, 2016, 91(6): 1228-1243.



国内最大病毒载体现货库
青年学术咖交流研发中心