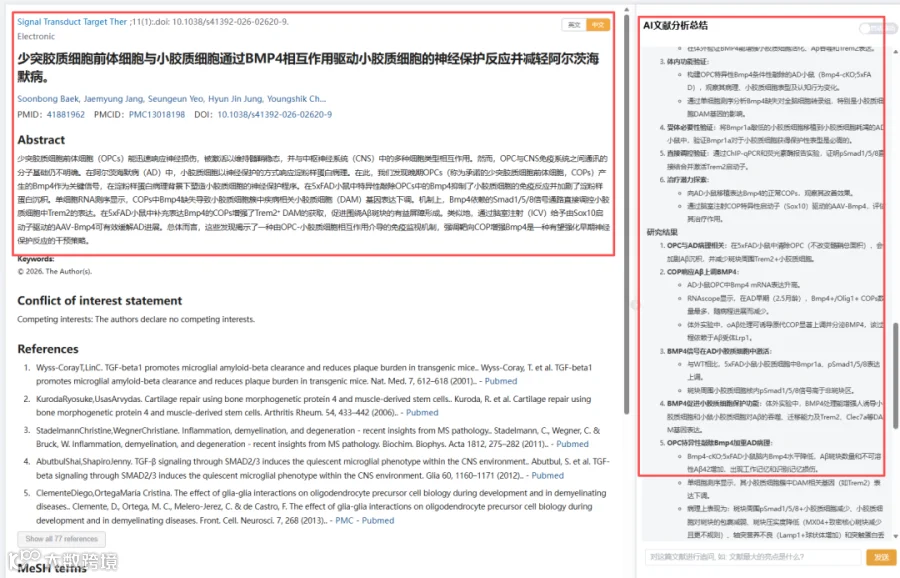
阿尔茨海默病(AD)的早期病理特征包括β淀粉样蛋白(Aβ)沉积和髓鞘损伤。少突胶质前体细胞(OPCs)是中枢神经系统的髓鞘修复细胞,但其在免疫调控中的角色尚不明确。这项发表于《Signal Transduction and Targeted Therapy》的研究,利用单细胞RNA测序(scRNA-Seq)、基因敲除小鼠、细胞移植及AAV基因治疗等多种技术,首次揭示:在AD病理刺激下,晚期OPCs(即“定型少突胶质前体细胞”,COPs)高表达并分泌BMP4。BMP4作用于小胶质细胞,通过Smad1/5/8信号通路直接结合TREM2启动子,诱导TREM2表达,驱动小胶质细胞向“疾病相关小胶质细胞(DAM)”转化,形成包裹Aβ斑块的保护性屏障,抑制斑块扩散和神经元损伤。OPC中特异性敲除Bmp4导致小胶质细胞功能缺陷、斑块增大、突触丢失和认知下降。而移植Bmp4表达的COPs或脑室内注射AAV-Sox10-Bmp4可显著改善AD病理和记忆功能。该研究为AD早期干预提供了全新细胞靶点和治疗策略。
今天给大家解读一篇3月发表在《Signal Transduction and Targeted Therapy》上的题目为“Oligodendrocyte precursor cells-microglia crosstalk via BMP4 drives microglial neuroprotective response and mitigates Alzheimer's disease.”的文章。本研究旨在探究OPC与中枢神经系统免疫系统交流的分子基础,特别是在AD中的作用。通过单细胞测序、条件性基因敲除、细胞移植、行为学测试等多种技术手段,作者发现AD模型(5xFAD)小鼠中,COP高表达BMP4。COP来源的BMP4作用于小胶质细胞上的Bmpr1a受体,激活Smad1/5/8信号,直接促进Trem2转录。这一信号轴对于小胶质细胞向保护性DAM状态转变、聚集并压实Aβ斑块至关重要。阻断该通路会加剧Aβ沉积和认知损伤,而增强该通路(如移植正常COPs或脑室注射AAV-Sox10-Bmp4)则能促进保护性小胶质细胞反应,减轻病理并改善记忆。研究确立了一条新的OPC-小胶质细胞通讯通路,为AD的早期治疗干预提供了新靶点。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!


团队成员合影(位于上海陆家嘴中心,可随时预约参观)
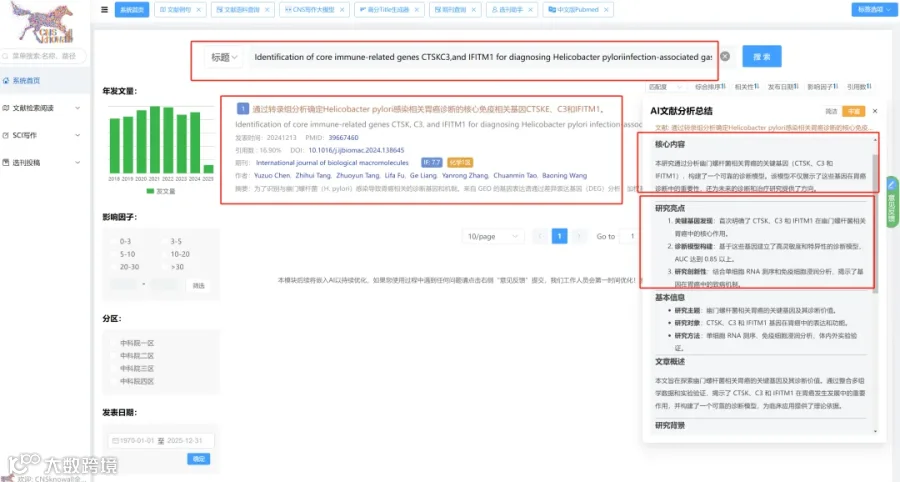
题目:《少突胶质细胞前体细胞与小胶质细胞通过BMP4相互作用驱动小胶质细胞的神经保护反应并减轻阿尔茨海默病》Oligodendrocyte precursor cells-microglia crosstalk via BMP4 drives microglial neuroprotective response and mitigates Alzheimer's disease
发表期刊:Signal Transduction and Targeted Therapy
影响因子:52.7
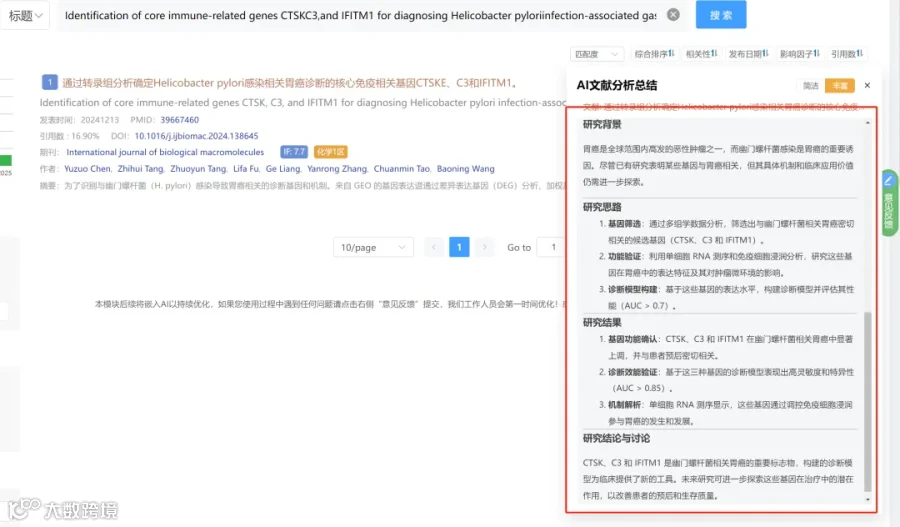
研究背景:
-
AD以Aβ和tau病理积累导致的进行性神经退行为特征,常伴随髓鞘完整性的广泛破坏。
-
除了作为祖细胞的经典角色,OPC已成为中枢神经系统稳态的多功能调节者,参与免疫调节等多种过程。
-
单细胞转录组学将OPC谱系进展细分为多个中间状态,其中COPs是转录上准备髓鞘化的晚期OPC群体,高表达Bcas1、Bmp4等标记物。
-
在AD中,小胶质细胞对Aβ挑战的早期反应会驱动其向具有神经保护功能的DAM表型转变,该表型以TREM2高表达为特征,能重塑淀粉样斑块。
-
BMP4在多种生物系统中与免疫调节和 tissue 稳态相关,且在AD患者脑脊液中水平升高。
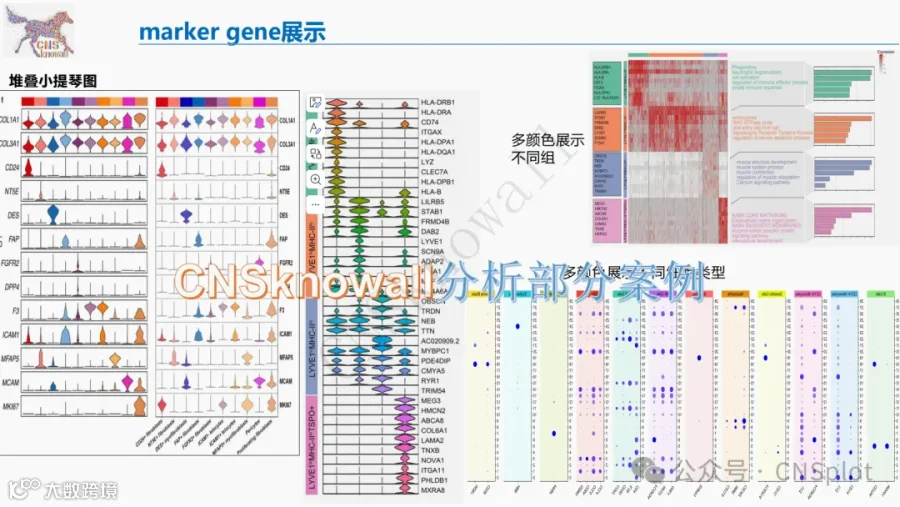
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
- 现象观察
发现在AD小鼠模型中,清除OPC会加重Aβ负担并减少小胶质细胞的Trem2表达,且COP高表达Bmp4。
- 分子机制探究
-
-
在AD模型小鼠和患者脑中,确认小胶质细胞Bmpr1a及其下游pSmad1/5/8信号激活。
-
在体外验证BMP4能增强小胶质细胞活化、Aβ吞噬和Trem2表达。
- 体内功能验证
-
构建OPC特异性Bmp4条件性敲除的AD小鼠(Bmp4-cKO;5xFAD),观察其病理、小胶质细胞表型及认知行为变化。
-
通过单细胞测序分析Bmp4缺失对全脑细胞转录组,特别是小胶质细胞DAM基因的影响。
- 受体必要性验证
将Bmpr1a敲低的小胶质细胞移植到小胶质细胞耗竭的AD小鼠中,验证Bmpr1a对于小胶质细胞获得保护性表型是必需的。
- 直接调控验证
通过ChIP-qPCR和荧光素酶报告实验,证明pSmad1/5/8直接结合并激活Trem2启动子。
- 治疗潜力探索
-
向AD小鼠移植表达Bmp4的正常COPs,观察其改善效果。
-
通过脑室注射COP特异性启动子(Sox10)驱动的AAV-Bmp4,评估其治疗作用。
研究亮点:
- 细胞互作新发现
首次揭示了OPC(特别是COP)与小胶质细胞之间通过BMP4-Bmpr1a轴进行交流,是AD早期神经免疫监测的关键机制。
- 关键信号分子
确定了COP是AD脑中BMP4的关键细胞来源,且其表达受Aβ寡聚体(oAβ)上调。
- 直接转录调控
阐明了BMP4下游的pSmad1/5/8能够直接结合并激活Trem2基因的启动子,将细胞间信号与核心AD保护基因直接联系起来。
- 双向功能验证
通过条件性敲除COP中的Bmp4加重病理,以及补充表达BMP4的COPs或给予AAV-Bmp4治疗减轻病理,双向证明了该通路的功能重要性。
- 治疗潜力
提出并验证了靶向COP增强BMP4表达(如AAV-Sox10-Bmp4)可作为早期干预AD、增强神经保护反应的潜在策略。
研究结果:
- OPC与AD病理相关
在5xFAD小鼠中清除OPC(不改变髓鞘总面积),会加剧Aβ沉积,并减少斑块周围Trem2+小胶质细胞。
- COP响应Aβ上调BMP4
-
-
RNAscope显示,在AD早期(2.5月龄),Bmp4+/Olig1+ COPs数量最多,随病程进展而减少。
-
体外实验中,oAβ处理可诱导原代COP显著上调并分泌BMP4,该过程依赖于Aβ受体Lrp1。
- BMP4信号在AD小胶质细胞中激活
-
与WT相比,5xFAD小鼠小胶质细胞中Bmpr1a、pSmad1/5/8表达上调。
-
斑块周围小胶质细胞核内pSmad1/5/8信号高于非斑块区。
- BMP4促进小胶质细胞保护功能
体外实验中,BMP4处理能增强人诱导小胶质细胞和小鼠小胶质细胞对Aβ的吞噬、迁移能力及Trem2、Clec7a等DAM基因表达。
- OPC特异性敲除Bmp4加重AD病理
-
Bmp4-cKO;5xFAD小鼠脑内Bmp4水平降低,Aβ斑块数量和不可溶性Aβ42增加,出现工作记忆和识别记忆损伤。
-
单细胞测序显示,其小胶质细胞簇中DAM相关基因(如Trem2)表达下调。
-
病理上表现为:斑块周围pSmad1/5/8+小胶质细胞减少、小胶质细胞对斑块的包裹减弱、斑块压实度降低(MX04+致密核心斑块减少且更不规则)、轴突营养不良(Lamp1+球状体增加)和突触蛋白丢失。
- Bmpr1a是小胶质细胞响应BMP4所必需
-
敲低BMPR1A会抑制hiMG中BMP4诱导的TREM2上调。
-
将Bmpr1a敲低的小胶质细胞移植到AD小鼠中,移植细胞无法有效激活pSmad1/5/8信号、获得Trem2+ DAM表型,也不能有效压实斑块。
- pSmad1/5/8直接调控Trem2转录
-
ChIP-qPCR显示,在AD小鼠脑组织中,pSmad1/5/8(而非pSmad2/3)与Trem2启动子区域结合显著富集。
-
荧光素酶报告实验证实,BMP4能增强Trem2启动子活性。
- 增强BMP4通路具有治疗潜力
-
向AD小鼠移植正常COPs(而非Bmp4 KO的COPs),能增加斑块周围Trem2+小胶质细胞,减少Aβ斑块,并保护突触前结构。
-
脑室注射AAV-Sox10-Bmp4(COP特异性表达),能提高脑内Bmp4水平,增加Trem2+小胶质细胞,促进斑块压实,减少轴突病变和突触丢失,并显著改善小鼠的Y迷宫和新物体识别记忆。
研究总结:
- 主要结论
本研究确立了一条新的COP-小胶质细胞通讯轴,其中COP来源的BMP4作为指导性信号,通过激活小胶质细胞Bmpr1a/pSmad1/5/8通路直接上调Trem2,驱动小胶质细胞获得早期神经保护性DAM表型,从而限制Aβ病理进展。
- OPC功能多样性
研究拓展了对OPC功能的认识,表明COPs不仅是髓鞘修复的前体细胞,还是通过分泌BMP4调节中枢神经系统免疫环境的关键参与者。
- 与TGF-β信号的比较
尽管TGF-β信号也对小胶质细胞功能很重要,但本研究明确指出在AD背景下,BMP4-pSmad1/5/8通路对Trem2的调控具有特异性且至关重要,提示不同情境下信号的精确分工。
- 治疗意义
靶向COP并增强其BMP4表达(如AAV-Sox10-Bmp4),是一种有前途的干预策略。它旨在增强AD早期内源性的神经保护免疫反应,可能延长疾病的临床前阶段。文章也指出,BMP4可能同时作用于神经元、星形胶质细胞等其他脑细胞,共同营造神经保护环境。
- 局限性/未来方向
文章提到,由于COPs是短暂的前体细胞,其持久作用可能有限。此外,OPC本身具有的吞噬活性等其他功能在AD中的作用,值得进一步研究。
结果译文:
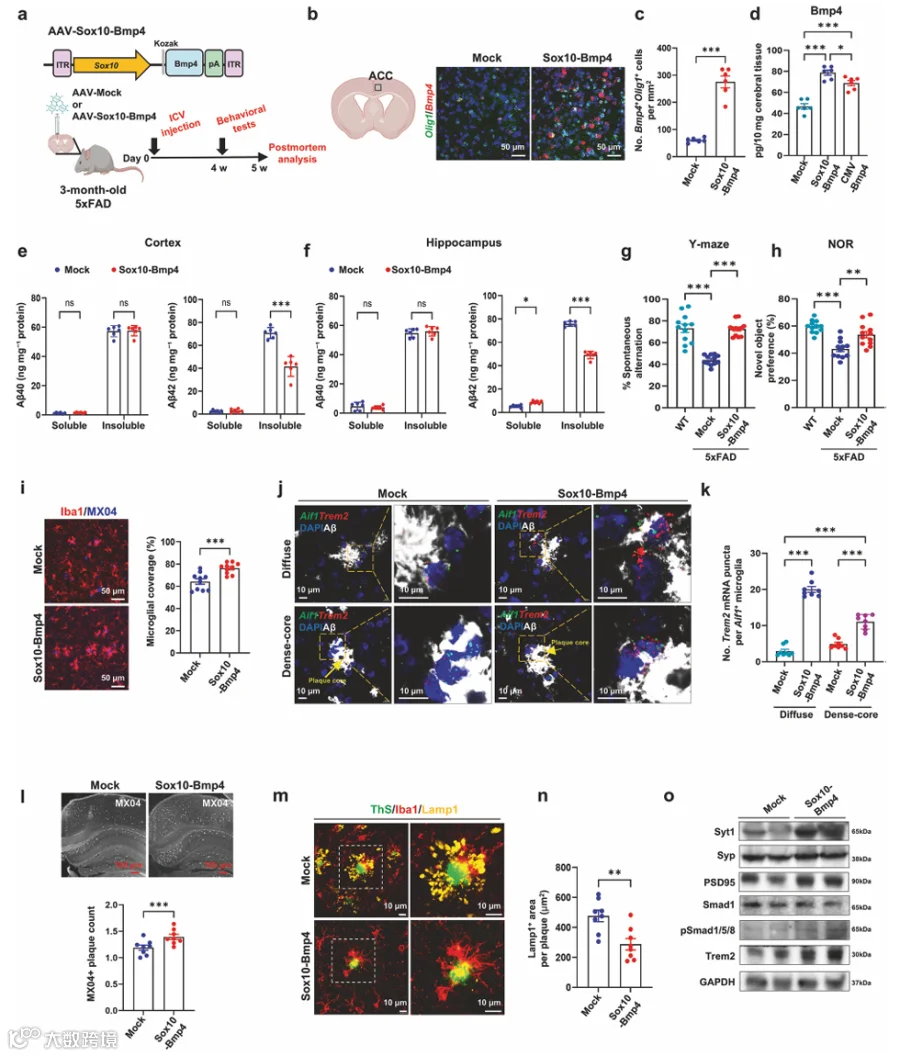
1.OPCs的消融独立于其在髓鞘维持中的作用而加剧Aβ负荷
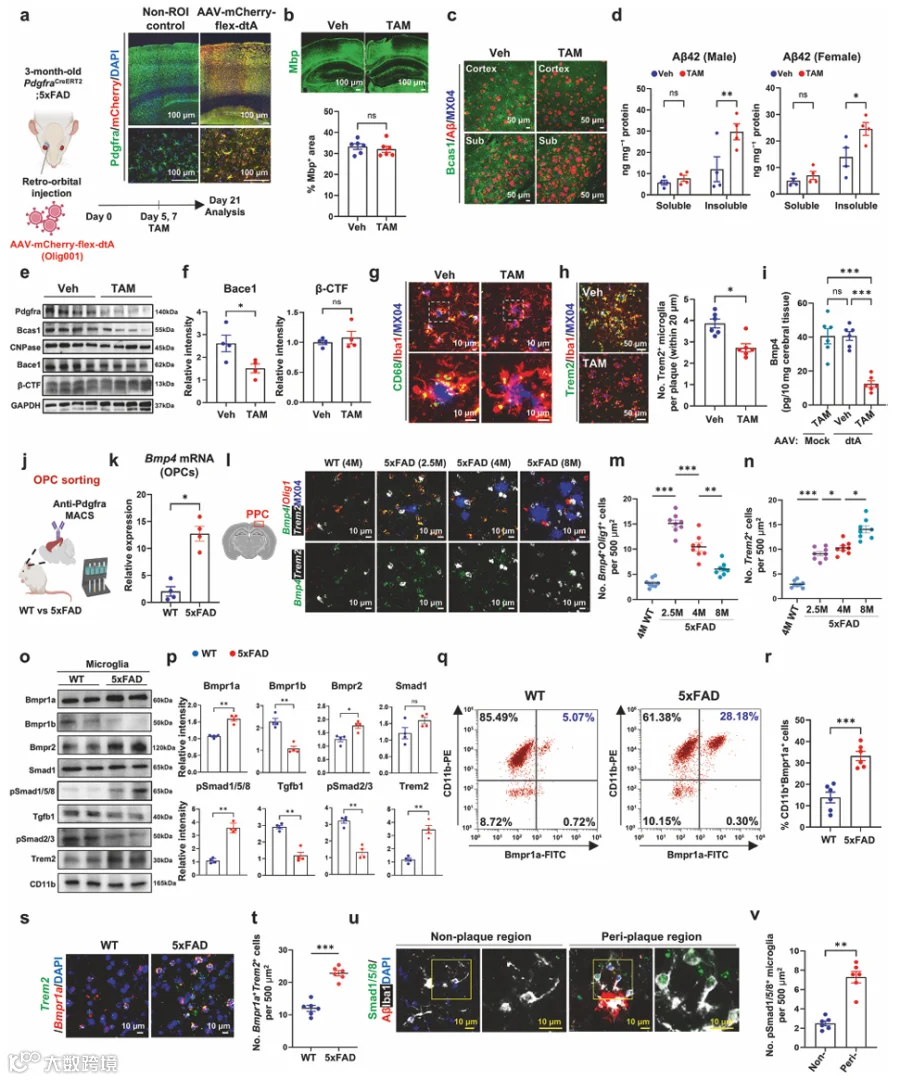
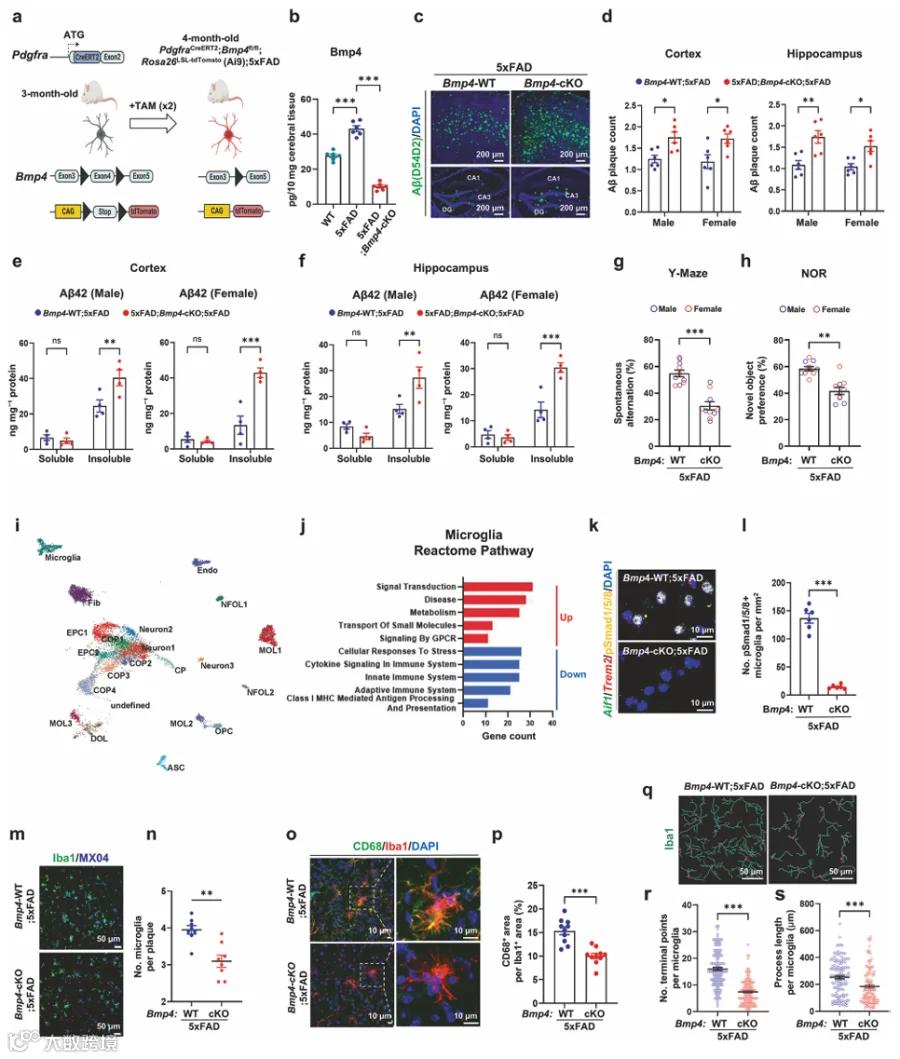
为了测试OPCs是否独立于维持髓鞘完整性的作用而参与淀粉样病理,我们将AAV-mCherry-flex-dTA经ROI注射到3月龄PdgfraCreERT2;5xFAD小鼠中(图1a)。我们证实了大多数Pdgfra⁺ OPCs被AAV-mCherry-flex-dTA成功转导(图1a,补充图1a),整体髓鞘化面积没有可检测的变化(图1b)。Bcas1⁺ COPs是AD小鼠中髓鞘修复的关键群体。TAM给药使注射AAV-mCherry-flex-dTA的5xFAD小鼠中Bcas1⁺ COPs减少了约70-80%(补充图1b、c)。值得注意的是,dtA介导的COPs消融导致皮质和Sub中淀粉样斑块计数显著增加(图1c,补充图1d)。ELISA进一步证实不溶性组分中的Aβ42水平显著升高(图1d),而Aβ40水平保持不变(补充图1e)。与我们的免疫标记结果一致,OPC标志物Pdgfra和COP标志物Bcas1减少,而成熟OL标志物CNPase的表达保持不变(图1e,补充图1f)。OPC消融后Bace1蛋白水平降低(图1e、f),支持OPCs表达Bace1的观点。尽管Bace1蛋白水平降低,β-CTF保持不变(图1e、f)。我们发现OPC消融导致小胶质细胞中CD68⁺吞噬体减少(图1g,补充图1g)。此外,OPC消融显著减少了4月龄5xFAD小鼠中Trem2⁺小胶质细胞的数量(图1h)。
我们对已发表的PS2APP模型单细胞RNA测序数据集(GSE160512)中OL谱系细胞的转录谱进行了UMAP聚类分析(补充图2a),显示COP簇表现出典型COP标志物的高表达(补充图2b)。与P301L和CPZ模型相比,PS2APP小鼠的COPs以Bmp4表达升高为特征(补充图2c)。由dtA介导的Pdgfra⁺ OPCs消融导致TBS可溶性脑裂解物中Bmp4蛋白水平降低(图1i),促使我们研究COP来源的Bmp4在抗淀粉样病理的CNS免疫应答中的功能。
2.在淀粉样蛋白应答中,COPs上调Bmp4的表达和分泌
抗Pdgfra磁珠分选OPCs的qRT-PCR分析(图1j)显示Pdgfra高表达,而神经元标志物NeuN不存在(补充图2d)。我们观察到与WT小鼠相比,5xFAD小鼠分选出的OPCs中Bmp4 mRNA表达增加(图1k)。接下来,我们使用RNAscope-ISH分析了PPC中Bmp4⁺ COP动态与AD进展的关系。Bmp4⁺/Olig1⁺ COPs在2.5月龄5xFAD小鼠中最为丰富(图1l、m)。随着AD进展,Trem2⁺小胶质细胞逐渐增加,而Bmp4⁺/Olig1⁺ COPs群体逐渐减少(图1m、n)。在8月龄时,5xFAD小鼠表现出明显的神经炎症,其特征是广泛的Aβ斑块沉积(补充图2e)和炎症性Trem2⁺小胶质细胞的大量增加(图1n)。Bmp4⁺ COPs在弥漫性淀粉样斑块附近不存在,但在致密核心斑块周围定位(补充图2f)。对AD患者死后组织的IHC分析显示,与非斑块区域中少数BMP4和BCAS1免疫反应性细胞相比,在斑块周围区域经常观察到表达BMP4的BCAS1⁺细胞(补充图2g、h)。对已发表scRNA-seq数据集的分析显示,Bmp4表达在COP群体中高度富集(补充图2i-k,补充表1)。5xFAD小鼠大脑的IHC显示,Bmp4在COPs中选择性表达,在星形胶质细胞、小胶质细胞、神经元或神经干细胞中未检测到表达(补充图2l、m)。
为了评估Aβ是否影响COP中Bmp4的表达,我们将OPCs分化为原代COPs(补充图3a)。暴露于寡聚Aβ诱导Bmp4 mRNA显著上调(补充图3b)。RNAscope-ISH验证了这一上调,显示oAβ处理后原代COPs中Bmp4 mRNA斑点显著增加(补充图3c、d)。与此一致,oAβ处理显著提高了Bmp4蛋白水平(补充图3e、f)。在各种原代脑细胞类型中,COPs对oAβ表现出最高的Bmp4表达水平(补充图3g)。此外,Aβ暴露导致条件培养基以及外泌体中Bmp4水平显著增加(补充图3h、i)。值得注意的是,与CM中的可溶性部分相比,通过外泌体释放的Bmp4比例显著增加(补充图3j、k)。已知Lrp1可跨血脑屏障转运Aβ,并在OPCs中高表达。PLA显示,在原代COPs中Aβ与Lrp1直接相互作用(补充图3l、m)。用针对先前验证的靶序列的shLrp1转染,显著减少了原代COPs中oAβ的内化(补充图3l-o)。Lrp1敲低减弱了Aβ诱导的Bmp4 mRNA水平上调(补充图3p)。
3.Smad1/5/8信号在表达Bmpr1a的斑块相关小胶质细胞中被激活
从4月龄5xFAD小鼠分离的小胶质细胞与年龄匹配的WT小鼠相比,表现出Tgfβ1及其下游效应物pSmad2/3的表达降低,同时Trem2表达上调(图1o、p)。这些变化表明小胶质细胞静息状态特征的丧失和向疾病相关状态的转变。有趣的是,Bmpr1a及其下游效应物pSmad1/5/8的表达在5xFAD小鼠中上调(图1o、p)。Bmp2表达也增加(图1o、p),而Bmpr1b蛋白在5xFAD小鼠中的表达水平显著低于Bmpr1a(补充图4a)。CD11b富集小胶质细胞的流式细胞术分析显示,与WT小鼠相比,5xFAD小鼠的小胶质细胞表达显著更高水平的Bmpr1a(图1q、r)。我们还在5xFAD小鼠皮质中观察到共表达Bmpr1a的Trem2⁺小胶质细胞增加(图1s、t)。在Aβ斑块附近观察到pSmad1/5/8⁺小胶质细胞的数量高于非斑块区域(图1u、v)。同样,与非痴呆对照受试者相比,人类AD受试者的IBA1⁺小胶质细胞中pSmad1/5/8水平也增加(补充图4b、c)。
4.体外分析显示Bmp4促进小胶质细胞活化,增强Aβ清除
为了评估Bmp4对小胶质细胞活化状态的影响,我们生成了人诱导小胶质细胞(补充图4d)。rhBMP4处理导致pSMAD1/5/8以及TREM2上调(补充图4e、f)。在hiMG中,rhBMP4增强了对pHrodo标记的Aβ的吞噬作用,并上调了DAM的关键标志物CLEC7A,而rhTGFβ1则抑制Aβ吞噬(补充图4g-j)。rhBMP4诱导的小胶质细胞吞噬反应增强被Bmp受体抑制剂LDN-193189共同处理所消除(补充图4h-j)。
接下来,我们进一步分析了小鼠小胶质细胞中oAβ响应的功能变化。Transwell迁移实验显示,经oAβ处理的BV2细胞在rmBMP4处理后表现出显著增加的迁移能力(补充图4k、l)。在来自过表达Bmp4的原代COP的条件培养基中培养的原代小胶质细胞,其pSmad1/5/8的核定位和Clec7a表达增加,这一效应被LDN-193189逆转(补充图4m-p)。qRT-PCR分析显示,单独的Bmp4足以在oAβ处理的原代小胶质细胞中诱导Trem2表达(补充图4q)。值得注意的是,rmBMP4和rmIFN-γ的共同处理导致DAM基因如Clec7a和Clec2d以及IFN应答基因Stat1、Stat2和Irf7的上调(补充图4q)。这些数据表明,Bmp4诱导Trem2表达是IFN通路依赖的小胶质细胞向早期保护性表型(类似于早期DAM)转变的关键先决条件。
5.COPs中Bmp4的基因消融短暂恢复髓鞘完整性但加剧淀粉样蛋白负荷
为了精确研究COP来源的Bmp4在淀粉样病理进展中的作用,我们通过杂交产生了诱导性OPC特异性Bmp4条件性敲除;5xFAD小鼠,并与/不与Rosa26-SL-tdTomato(Ai9)杂交(图2a)。在TAM给药后5天,约90%的Bcas1⁺ COPs强表达tdTomato(补充图5a、b)。TAM给药后5天,分选的Pdgfra⁺ OPCs中Bmp4 mRNA表达基本被消除(补充图5c)。在Bmp4-cKO;5xFAD小鼠中,Aβ斑块周围的tdTomato⁺ OPCs数量与Bmp4-WT;5xFAD小鼠相比无显著差异(补充图5d)。ELISA显示,在5xFAD小鼠中观察到的Bmp4水平升高在OPC特异性Bmp4-cKO中被基本消除(图2b)。值得注意的是,Bmp4-cKO导致Aβ斑块的大小和数量显著增加(补充图5e,图2c、d)。接下来,我们通过ELISA测量了Aβ种类的浓度。尽管可溶性Aβ40、不溶性Aβ40和可溶性Aβ42水平没有变化,但我们在Bmp4-cKO;5xFAD中观察到不溶性Aβ42显著增加(补充图5f、g,图2e、f)。OPC中Bmp4的基因消融通过Y迷宫和NOR测试评估,损害了工作记忆和学习记忆(图2g、h)。
接下来,我们对4月龄Bmp4-WT;5xFAD和年龄匹配的Bmp4-cKO;5xFAD的皮质细胞进行了scRNA-seq。UMAP分析识别出21种不同的细胞类型(图2i),单个簇表现出特征性标志物的表达(补充图6a,补充表2)。DEGs比较显示,OL谱系亚型的转录组因Bmp4-cKO而改变(补充图6b,补充表3)。OPC和COP3簇的DEGs包括大量Smad4靶基因(补充图6c)。OPC簇中上调的基因参与蛋白定位到质膜、磷酸离子转运和膜蛋白水解(补充图6d,补充表4)。此外,下调的基因与BMP信号通路的调控相关(补充图6d)。
COP3簇的比例因OPC特异性Bmp4缺失而大大减少(6.2% → 1.2%,补充图6e)。火山图中的DEGs显示,在Bmp4-cKO;5xFAD的COP3簇中,Plp1和Mbp显著上调(补充图6f)。对从Bmp4-cKO;5xFAD分选的抗O4 MACS OLs进行的qRT-PCR分析证实,成熟OL标志物Cnp、Mbp和Apc上调,而Bcas1下调(补充图6g)。在COP3簇内,GO分析显示上调基因在细胞对Aβ的反应和髓鞘维持相关通路中优先富集(补充图6h,补充表4)。最近的一项研究报道,髓鞘完整性的丧失是AD病理启动的上游因素,且髓鞘的基因缺失可减轻Aβ沉积。在4月龄时(即TAM给药后1个月),Bmp4-cKO;5xFAD皮质区域的Mbp IHC显示髓鞘面积增加(补充图6i)。然而,到7月龄时,皮质中Aβ和Mbp的免疫标记显示,与Bmp4-WT;5xFAD相比,Bmp4-cKO;5xFAD中斑块数量显著增加(补充图6j),且Aβ斑块周围局灶性脱髓鞘增加(补充图6k)。
6.OPCs中Bmp4缺乏导致Aβ相关小胶质细胞功能反应减弱
在Bmp4-cKO;5xFAD小胶质细胞中,表达降低的基因富集于与细胞对应激的反应、细胞因子信号传导以及先天性和适应性免疫系统相关的通路(图2j,补充表5)。RNAscope-ISH显示,在Bmp4-cKO;5xFAD中Aβ斑块周围的Bmp4⁺ COPs消失(补充图7a、b)。OPC特异性Bmp4缺失后,Trem2⁺小胶质细胞中pSmad1/5/8的主要核分布基本消失(图2k、l)。小胶质细胞在Aβ斑块形成中具有多方面作用,首先清除Aβ,随后压缩和包裹斑块。我们发现OPC中Bmp4-cKO导致Aβ斑块周围的小胶质细胞聚集减少(图2m、n,补充图7c-e)。此外,吞噬性小胶质细胞中的CD68⁺吞噬体面积显著减少(图2o、p)。TAM给药后,Iba1⁺小胶质细胞显示出显著更少的末端分支点和更短的过程(图2q-s)。因此,OPC来源的Bmp4缺失导致小胶质细胞过程复杂性降低,与小胶质细胞监视和早期活化状态受损一致。
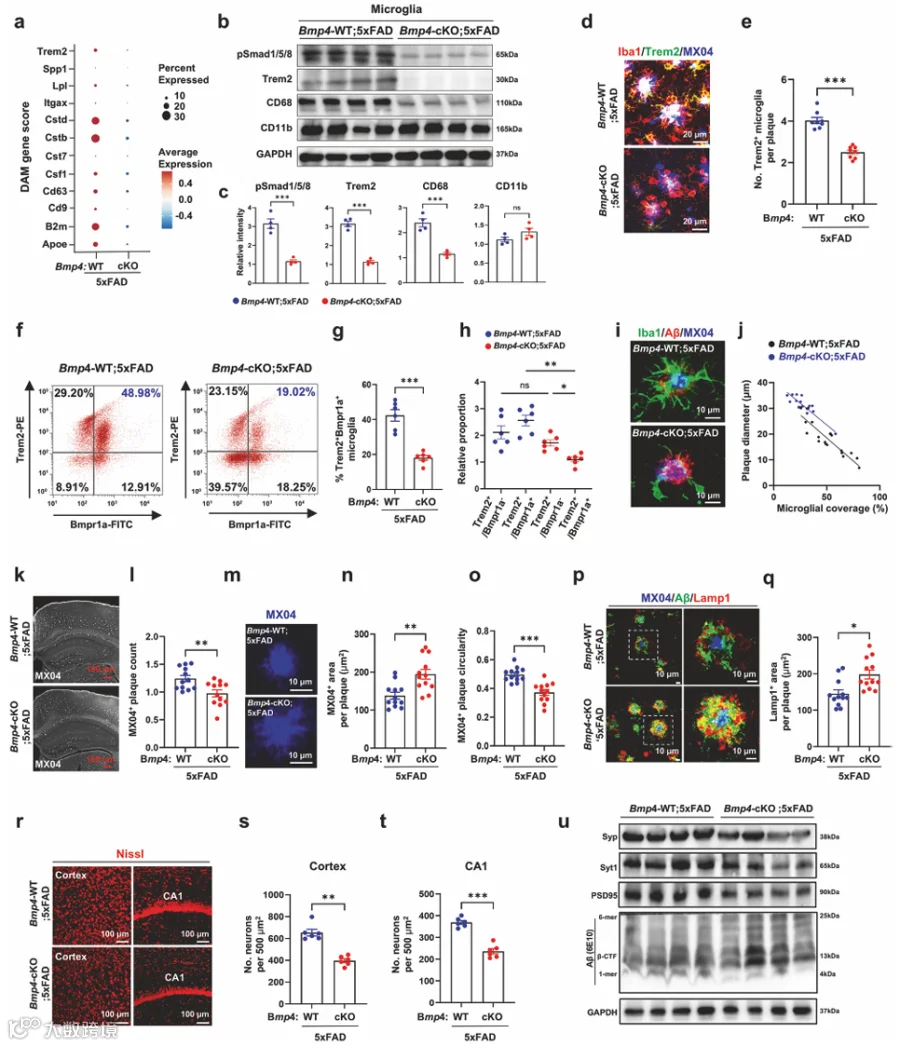
7.COP来源的Bmp4促进Trem2驱动的小胶质细胞屏障功能
在小胶质细胞簇的单细胞转录组中,DAM标志物Trem2、B2m、Ctsd和Ctsb在Bmp4-cKO;5xFAD中显著下调(图3a)。DAM基因表达降低(补充图7f、g)通过免疫印迹分析得到证实(图3b、c)。OPC特异性Bmp4缺失损害了小胶质细胞中Trem2下游信号传导,如pSyk减少所证明,这反过来抑制了Akt和mTOR通路的激活(补充图7h、i)。Bmp4-cKO使斑块相关区域中Trem2⁺小胶质细胞的数量大幅减少(图3d、e,补充图7j-l)。CD11b富集的小胶质细胞显示,与Bmpr1a阴性的Trem2⁺小胶质细胞相比,Bmp4-cKO小鼠中表达Bmpr1a的Trem2⁺小胶质细胞减少了约两倍(图3f-h)。免疫染色显示,Iba1⁺细胞中不表达外周巨噬细胞标志物CD163,而所有Iba1⁺细胞均表达小胶质细胞特异性标志物Trem119(补充图7m、n)。在未灌注的脑中检测到CD11b高/CD45高外周巨噬细胞,但在PBS灌注后经密度梯度血细胞去除的小胶质细胞中不存在(补充图7o、p)。因此,这些数据表明外周免疫细胞浸润不导致4月龄5xFAD小鼠中观察到的小胶质细胞表型。此外,Bmp4-cKO;5xFAD中DAM获得的缺失并非由于小胶质细胞凋亡(补充图7q、r)。
Bmp4-cKO使小胶质细胞突起对致密核心斑块的覆盖减少(图3i、j)。这种小胶质细胞屏障的缺失与TAM给药后MX04染色的致密核心斑块存在减少相一致(图3k、l)。在Bmp4-cKO;5xFAD中,MX04⁺斑块核心增大,但显示出显著降低的圆度,表明与Bmp4-WT;5xFAD相比压缩性更差(图3m-o)。scRNA-seq显示,Bmp4-cKO;5xFAD中Neuron1簇的下调基因富集于与神经生长因子反应、突触囊泡运输和神经元突触可塑性调节相关的通路(补充图8a和补充表4)。Neuron1的比例在Bmp4-cKO;5xFAD中增加(8.3% → 15.4%,补充图8b),可能表明与突触损伤相关的神经元亚型增加。点图(补充图8c)和基因得分(补充图8d)显示Bmp4-cKO导致突触基因下调。在Bmp4-cKO;5xFAD中观察到Lamp1⁺球体的积累增加(图3p、q),与Syt1⁺轴突球体的显著重叠(补充图8e、f),以及NF-M⁺轴突肿胀增加(补充图8g、h)。此外,Bmp4-cKO使5xFAD小鼠皮质和CA1区中Nissl染色神经元的数量显著减少(图3r-t)。使用免疫印迹分析评估突触标志物和Aβ的蛋白丰度。虽然在Bmp4-cKO;5xFAD中检测到的单体Aβ水平低于Bmp4-WT;5xFAD,但oAβ在Bmp4-cKO;5xFAD中急剧增加(图3u,补充图8i)。突触前(Syp和Syt1)和突触后(PSD95)标志物的表达水平在Bmp4-cKO;5xFAD中显著降低(图3u,补充图8i)。
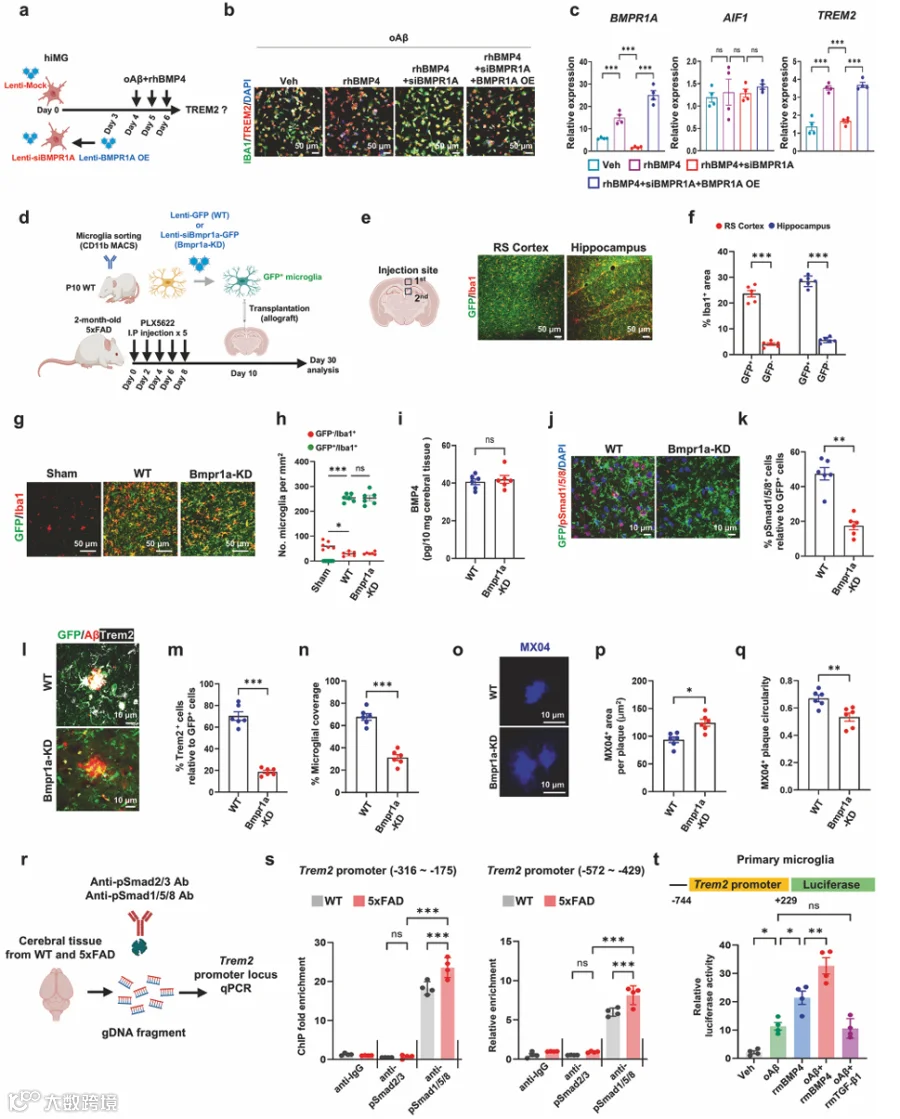
8.缺乏Bmpr1a的同种异体移植小胶质细胞无法获得Trem2相关的神经保护状态
hiMG中BMPR1A的siRNA介导的敲低显著减弱了oAβ和rhBMP4诱导的TREM2在蛋白(图4a、b,补充图9a)和mRNA水平(图4c)的上调,而不影响其他Bmp受体的表达(补充图9b)。这一效应通过BMPR1A过表达得到稳健恢复(图4b、c,补充图9a)。为了评估Bmp4是否通过Bmp受体激活小胶质细胞,我们将携带或缺乏Bmpr1a的小胶质细胞移植到5xFAD小鼠中。使用PLX5622建立急性小胶质细胞耗竭模型,每两天腹腔注射一次,共五次(图4d)。该方案有效耗竭了约70%的内源性小胶质细胞(补充图9c、d),允许评估移植小胶质细胞的功能。CD11b分选的小胶质细胞用编码GFP(WT)或siBmpr1a-GFP的慢病毒载体转导(图4d)。在测试的三个siRNA序列中,siRNA2在降低Bmpr1a表达方面效率最高,并用于所有后续分析(补充图9e)。将原代小胶质细胞立体定位注射到压后皮质区和海马区。尽管由于内源性小胶质细胞耗竭不完全和急性时间框架,移植的小胶质细胞没有广泛迁移,但移植的小胶质细胞在注射部位稳定植入(图4e、f)。WT和Bmpr1a-KD小胶质细胞均成功植入RS皮质(图4g、h)。与假手术对照相比,移植脑中GFP⁺小胶质细胞数量减少表明,移植的小胶质细胞抑制内源性小胶质细胞再生并有效整合到脑实质中(图4h)。ELISA显示,在移植WT或Bmpr1a-KD小胶质细胞的5xFAD小鼠中,皮质区域的总体Bmp4蛋白含量没有显著差异(图4i)。在植入位点的WT小胶质细胞中观察到强烈的pSmad1/5/8活化,而Bmpr1a-KD小胶质细胞未能激活该通路(图4j、k)。缺乏Bmpr1a的小胶质细胞在淀粉样斑块周围的Clec7a⁺小胶质细胞定位显著减少(补充图9f),并且在获得Trem2⁺ DAM方面表现出显著损伤(图4l、m)。此外,Bmpr1a-KD小胶质细胞未能有效地聚集在淀粉样斑块周围(图4n),导致在Bmpr1a-KD小胶质细胞移植区域中斑块压缩性降低(图4o-q)。
我们检查了Smad4与Trem2启动子的直接结合是否促进其转录激活。在BV2细胞中使用Smad4特异性抗体的ChIP-qPCR分析显示,覆盖Trem2启动子区域的DNA序列在Smad4-DNA复合物中富集(补充图9g)。用GFP或Smad4表达DNA构建体转染受Trem2启动子控制的荧光素酶报告基因到BV2细胞中,结果显示Smad4显著增加了荧光素酶活性(补充图9h)。由于Smad4是Bmp和Tgfb信号共享的下游效应物,我们使用针对通路特异性下游介质(Bmp信号的pSmad1/5/8和Tgfb信号的pSmad2/3)的抗体进行了体内ChIP-qPCR测定(图4r)。pSmad1/5/8显示出与Trem2启动子位点的强结合,且这种富集在5xFAD小鼠中比WT对照中更高。相比之下,pSmad2/3几乎不与TREM2启动子结合(图4s)。在原代小胶质细胞中,荧光素酶测定显示,rMTGF-β1和oAβ的共同处理未诱导TREM2启动子驱动的荧光素酶活性。相比之下,rBMP4和oAβ的共同处理导致荧光素酶活性显著增加(图4t)。
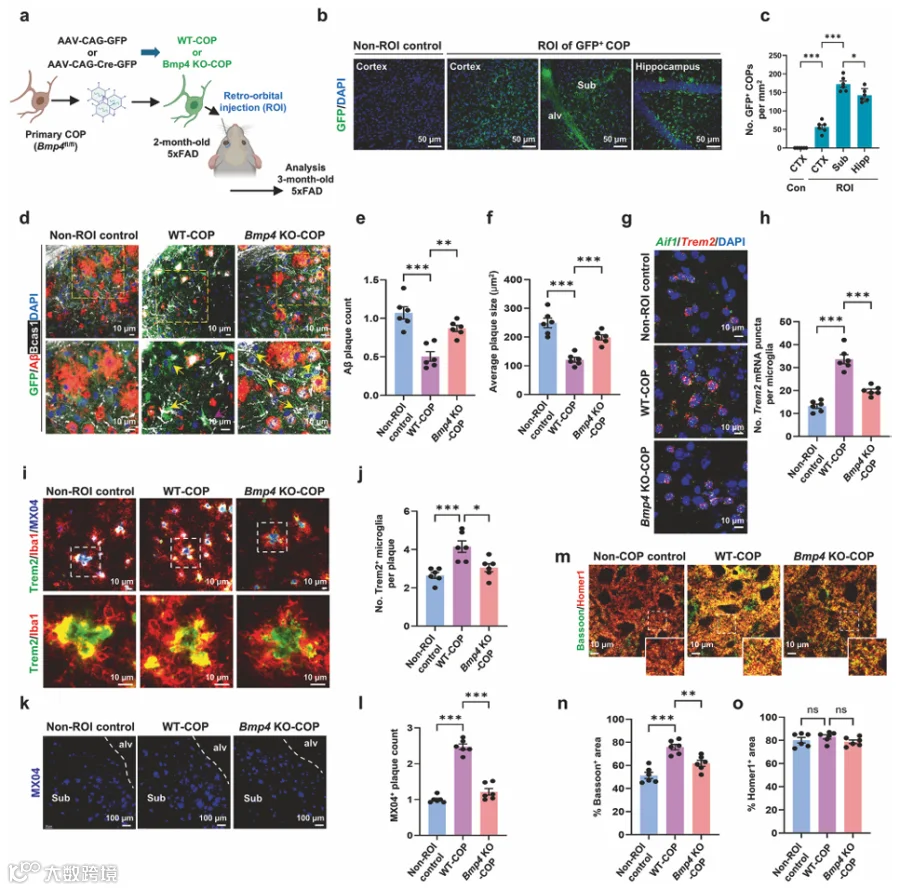
9.COPs的补充导致斑块相关TREM2⁺小胶质细胞增加
由于COPs是迅速分化为髓鞘化OLs的瞬时前体细胞,我们测试了移植额外的原代COPs是否可以补充内源性COPs并增强Bmp4介导的小胶质细胞活化。我们将来自Bmp4fl/fl小鼠并用AAV-CAG-GFP(WT-COP)或AAV-CAG-Cre-GFP(Bmp4 KO-COP)转导的原代COPs经ROI注射到2月龄5xFAD小鼠中(图5a)。使用原代培养物的体外转导显示,AAV-PHP.eb有效感染COPs,达到约90%的转导效率(补充图10a、b)。ROI后3周,组织学分析显示整体髓鞘化面积保持不变(补充图10c、d)。原代COPs主要定位于Sub和海马,在皮质中观察到最少的植入(图5b、c)。我们聚焦于Sub,该区域在3月龄5xFAD小鼠中具有显著的淀粉样斑块沉积。移植原代COPs后,表达Bmp4的Olig1⁺细胞显著增加(补充图10e、f)。移植WT-COPs后观察到pSmad1/5/8⁺小胶质细胞增加,而Bmp4 KO-COPs移植未再现这一反应(补充图10g、h)。在原代COP移植后,Sub中的Aβ斑块数量和大小的显著减少,在接受Bmp4 KO-COPs的ROI小鼠中未观察到(图5d-f)。我们发现Aif1⁺小胶质细胞中TREM2转录本显著升高(图5g、h),并且小胶质细胞通过表达TREM2的过程参与斑块压缩增加(图5i、j),这促进了MX04⁺致密核心斑块的发展(图5k、l)。尽管在此阶段突触后结构尚未受损,WT-COPs的ROI移植通过Bassoon染色证明保留了突触前完整性(图5m-o)。这些明显的表型变化在Bmp4 KO-COP移植小鼠中基本不存在(图5m-o)。
10.ICV递送AAV-Sox10-Bmp4增强小胶质细胞神经保护并减轻AD病理
我们设计了一个在Sox10启动子下表达Bmp4的AAV构建体,该启动子在COPs中驱动Bmp4表达。为了评估其体内效果,我们通过ICV注射将AAV-Sox10-Bmp4或对照AAV-Mock注入3月龄5xFAD小鼠(图6a)。侧脑室周围前扣带皮质的RNAScope-ISH显示,Bmp4⁺/Olig1⁺细胞显著增加(图6b、c)。ELISA进一步揭示,在AAV-Sox10-Bmp4注射后,CNS中Bmp4水平显著升高(图6d)。值得注意的是,尽管给予了等量的病毒滴度,但COP特异性AAV-Sox10-Bmp4产生的TBS可溶性Bmp4水平高于普遍表达的AAV-CMV-Bmp4(图6d)。ICV递送AAV-Sox10-Bmp4导致Aβ斑块的数量和大小显著减少(补充图11a-c)。ELISA显示,AAV-Sox10-Bmp4使不溶性Aβ42水平显著降低(图6e、f)。注射AAV-Sox10-Bmp4的5xFAD小鼠的海马依赖性短期空间工作记忆(图6g)和学习记忆(图6h)显著改善。
接下来,我们观察到,与注射AAV-Mock的小鼠相比,AAV-Sox10-Bmp4使皮质区域中pSmad1/5/8⁺小胶质细胞显著上调(补充图11d、e)。AAV-Sox10-Bmp4给药后CD68⁺小胶质细胞显著升高,显示Bmp4介导的吞噬活性增强(补充图11f、g)。AAV-Sox10-Bmp4给药后,Aβ斑块周围的小胶质细胞存在增加(图6i)。使用Trem2和Aif1探针的RNAscope-ISH结合MX04⁺ Aβ斑块的组织学检测显示,AAV-Sox10-Bmp4使弥漫性斑块和致密核心斑块周围的Trem2/Aif1⁺小胶质细胞均显著增加(图6j、k)。AAV-Sox10-Bmp4注射后观察到MX04⁺致密核心斑块显著增加(图6l),同时斑块面积减少和斑块圆度增加(补充图11h-j)。AAV-Sox10-Bmp4诱导小胶质细胞重塑,表现为突起延长和分支复杂性增加(补充图11k-m),表明对Aβ病理的反应性增强。
IHC显示,ICV注射AAV-Sox10-Bmp4使Lamp1⁺(图6m、n)和Syn1⁺(补充图11n、o)轴突球体显著减少。Western blot分析证实,在注射AAV-Sox10-Bmp4的小鼠的皮质组织中,pSmad1/5/8、Trem2和突触标志物(Syt1、Syp和PSD95)增加(图6o,补充图11p)。在注射后30天,4月龄AAV-Sox10-Bmp4和AAV-Mock注射的5xFAD小鼠之间的髓鞘化面积没有差异(补充图11q、r)。然而,到90 dpi时,在6月龄时,与AAV-Mock转导的对照相比,AAV-Sox10-Bmp4处理的5xFAD小鼠表现出年龄和淀粉样病理相关脱髓鞘的显著减轻(补充图11q、r)。
更多结果和补充图表:doi:10.1038/s41392-026-02620-9
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!