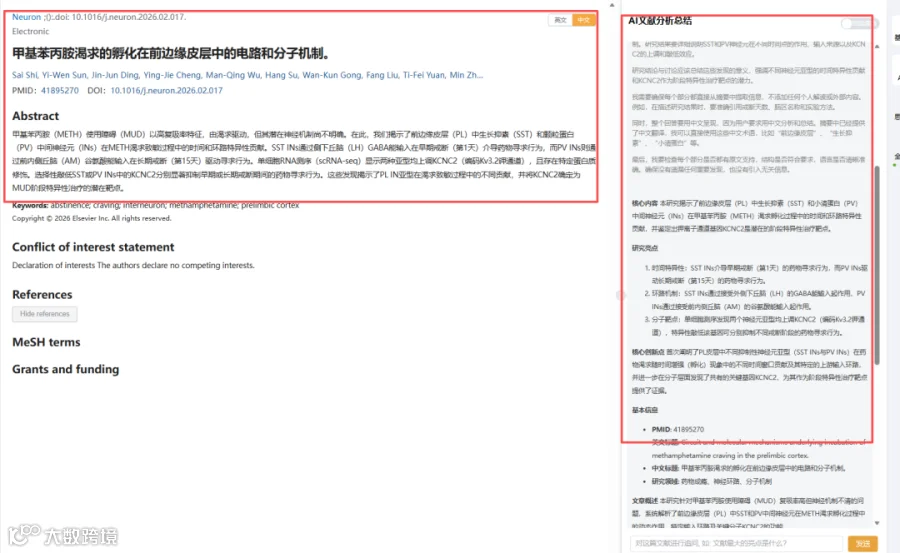
甲基苯丙胺成瘾的复吸率高达 80%,背后是“渴求孵育”这一核心机制在驱动。本研究利用单细胞转录组学与神经环路组学技术,首次揭示前边缘皮质中两类抑制性神经元在戒断不同阶段的“分工”奥秘:戒断早期,外侧下丘脑的 GABA 能输入抑制 SST 中间神经元,驱动渴求;戒断后期,丘脑前内侧的谷氨酸能输入激活 PV 中间神经元,维持渴求。单细胞测序进一步锁定 KCNC2(编码 Kv3.2 钾通道)在两类神经元中阶段特异性上调,精准敲低 KCNC2 可显著降低渴求行为。该研究为甲基苯丙胺成瘾的“阶段化治疗”提供了全新的分子与回路靶点。
今天给大家解读一篇3月发表在《Neuron》上的题目为“Circuit and molecular mechanisms underlying incubation of methamphetamine craving in the prelimbic cortex.”的文章。本研究针对甲基苯丙胺使用障碍(MUD)复吸率高但神经机制不清的问题,系统解析了前边缘皮层(PL)中SST和PV中间神经元在METH渴求孵化过程中的动态作用、特定输入环路及关键分子KCNC2的功能。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!


团队成员合影(位于上海陆家嘴中心,可随时预约参观)
题目:《甲基苯丙胺渴求的孵化在前边缘皮层中的电路和分子机制》Circuit and molecular mechanisms underlying incubation of methamphetamine craving in the prelimbic cortex
发表期刊:Neuron
影响因子:15
研究背景:
甲基苯丙胺使用障碍(MUD)以高复吸率为特征,由渴求驱动,但其潜在的神经机制尚不清楚。
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
-
探究PL皮层中SST和PV中间神经元在戒断不同阶段(第1天与第15天)对药物寻求行为的影响。
-
解析驱动这两种神经元亚型活动的特定上游神经输入来源。
-
利用单细胞RNA测序(scRNA-seq)寻找在两种神经元亚型中共同发生变化的分子。
-
通过在特定神经元亚型中敲低关键分子,验证其对不同阶段药物寻求行为的影响。
研究亮点:
-
时间特异性:SST INs介导早期戒断(第1天)的药物寻求行为,而PV INs驱动长期戒断(第15天)的药物寻求行为。
-
环路机制:SST INs通过接受外侧下丘脑(LH)的GABA能输入起作用,PV INs通过接受前内侧丘脑(AM)的谷氨酸能输入起作用。
-
分子靶点:单细胞测序发现两个神经元亚型均上调KCNC2(编码Kv3.2钾通道),特异性敲低该基因可分别抑制不同戒断阶段的药物寻求行为。
研究结果:
- 行为与环路
SST INs通过接受外侧下丘脑(LH)的GABA能输入,介导早期戒断(第1天)的药物寻求行为;而PV INs通过接受前内侧丘脑(AM)的谷氨酸能输入,驱动长期戒断(第15天)的药物寻求行为。
- 分子发现
scRNA-seq揭示SST和PV INs两个亚型均出现KCNC2(编码Kv3.2钾通道)的上调,并存在特定的蛋白质修饰。
- 功能验证
选择性敲低SST INs中的KCNC2能显著抑制早期戒断期的药物寻求;选择性敲低PV INs中的KCNC2则能显著抑制长期戒断期的药物寻求。
研究总结:
本研究揭示了PL皮层中间神经元亚型在渴求孵化过程中的不同贡献:SST INs和PV INs分别在早期和长期戒断阶段扮演关键角色,且由不同的上游脑区输入驱动。研究进一步发现并验证了KCNC2是这两个过程共有的关键分子,将其确定为MUD中具有阶段特异性的潜在治疗靶点。这些发现为理解渴求孵化的时空动态机制提供了新的视角。
结果译文:
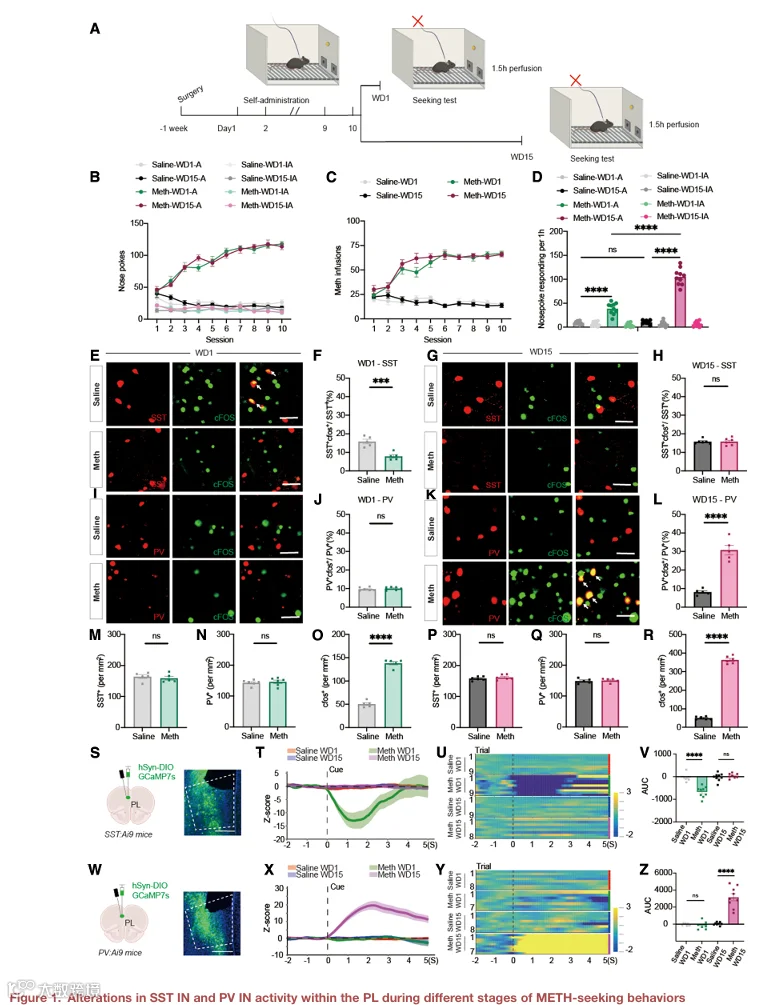
1.PL 内 SST IN 和 PV IN 在 METH 寻求行为不同阶段的活动变化
为了研究 PL 中不同中间神经元亚型在 METH 寻求行为中的作用,我们首先训练小鼠进行 METH 自身给药,随后在戒断第 1 天(WD1)或第 15 天(WD15)进行线索诱导的药物寻求测试(图 1A)。与盐水对照组相比,METH 训练组在 WD1 和 WD15 均表现出显著升高的主动鼻触次数,表明成功建立了线索诱导的 METH 寻求行为(图 S1A–S1C)。
为了确定哪些中间神经元亚型在 METH 寻求过程中被激活,我们在 WD1 和 WD15 进行线索诱导寻求测试后,通过免疫荧光染色检测了 PL 中 c‑Fos 与 SST、PV 或 VIP 的共定位(图 1B–1E)。在 WD1,c‑Fos⁺ 细胞中与 SST 共定位的比例显著增加(图 1F),而与 PV 或 VIP 共定位的比例无显著变化(图 1G–1J)。相反,在 WD15,c‑Fos⁺ 细胞中与 PV 共定位的比例显著升高(图 1H),而 SST 和 VIP 无显著变化(图 1F 和 1J)。这些结果表明,SST IN 在 WD1 被优先激活,而 PV IN 在 WD15 被优先激活。
为进一步验证这一发现,我们使用纤维光度法实时记录了 WD1 和 WD15 线索诱导寻求测试期间 PL 中 SST IN 和 PV IN 的钙活动(图 1K–1M)。在 WD1,SST IN 在测试期间表现出显著升高的钙信号,而 PV IN 无明显变化(图 1N–1Q)。在 WD15,PV IN 的钙活动显著增强,而 SST IN 无明显变化(图 1R–1U)。此外,我们还记录了不同戒断期谷氨酸能神经元的活动,发现其在 WD1 和 WD15 寻求测试中均被激活(图 S2I–S2L)。总之,这些结果表明不同中间神经元类型在线索诱导的寻求行为中可能发挥独特作用。
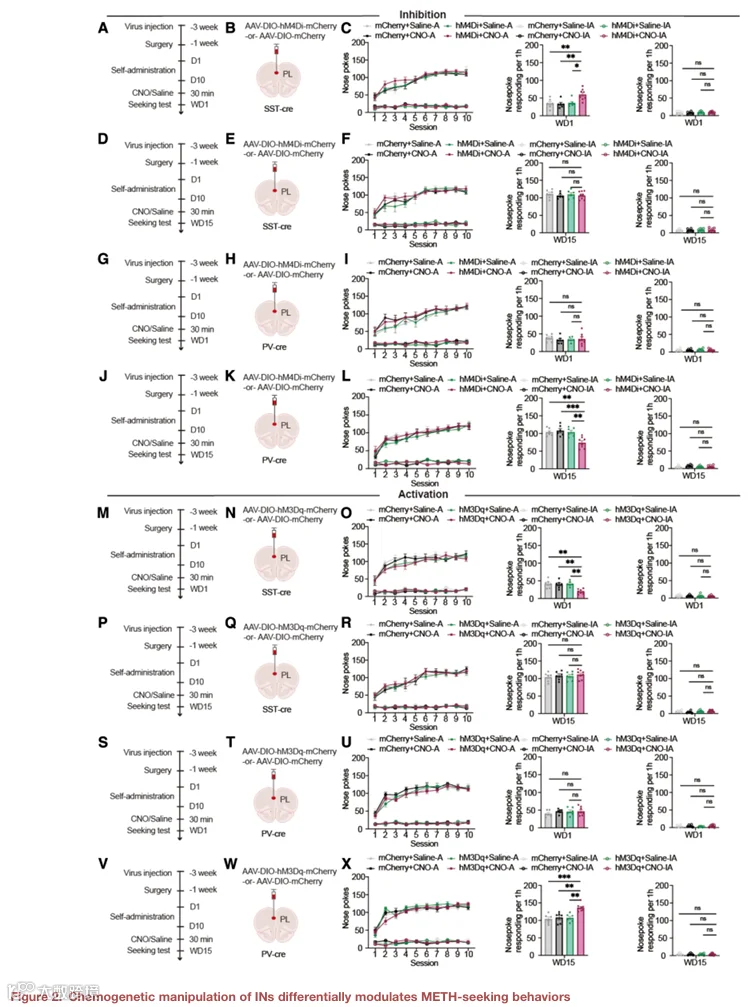
2.操纵 SST IN 和 PV IN 差异性地改变 METH 寻求行为的不同阶段
接下来,为了直接验证 SST IN 或 PV IN 在 METH 寻求行为中的必要性,我们采用了化学遗传学操纵。在 PL 内选择性表达 Cre 依赖的抑制性 DREADD(AAV‑DIO‑hM4Di‑mCherry 或 AAV‑DIO‑mCherry)。在行为测试前 30 分钟给予生理盐水或 CNO,以评估线索诱导的 METH 寻求行为。病毒注射位点、表达和效率首先得到验证(图 S4A–S4F)。结果显示,mCherry 表达小鼠和 hM4Di 表达小鼠均获得了稳定的 METH SA。CNO 介导的 SST IN 抑制在 WD1 寻求测试期间增加了主动鼻触次数(图 2A–2C),但这一效应在 WD15 无统计学意义(图 2D–2F)。相反,抑制 PV IN 对 WD1 寻求行为无影响(图 2G–2I),但在 WD15 显著减少了主动鼻触次数(图 2J–2L)。
此外,为了验证 SST IN 或 PV IN 在介导 METH 寻求行为中的充分性,我们还在 PL 中使用了 Cre 依赖的兴奋性 DREADD(AAV‑DIO‑hM3Dq‑mCherry 或 AAV‑DIO‑mCherry)。化学遗传学激活 SST IN 在 WD1 抑制了寻求行为(图 2M–2O),但在 WD15 无影响(图 2P–2R)。相反,激活 PV IN 对 WD1 无影响(图 2S–2U),但在 WD15 显著增加了寻求行为(图 2V–2X)。综上,这些发现表明 SST IN 主要在戒断早期调节 METH 寻求行为,而 PV IN 则在戒断后期影响 METH 寻求行为,提示这些中间神经元群体在 METH 渴求中具有不同的时间作用。
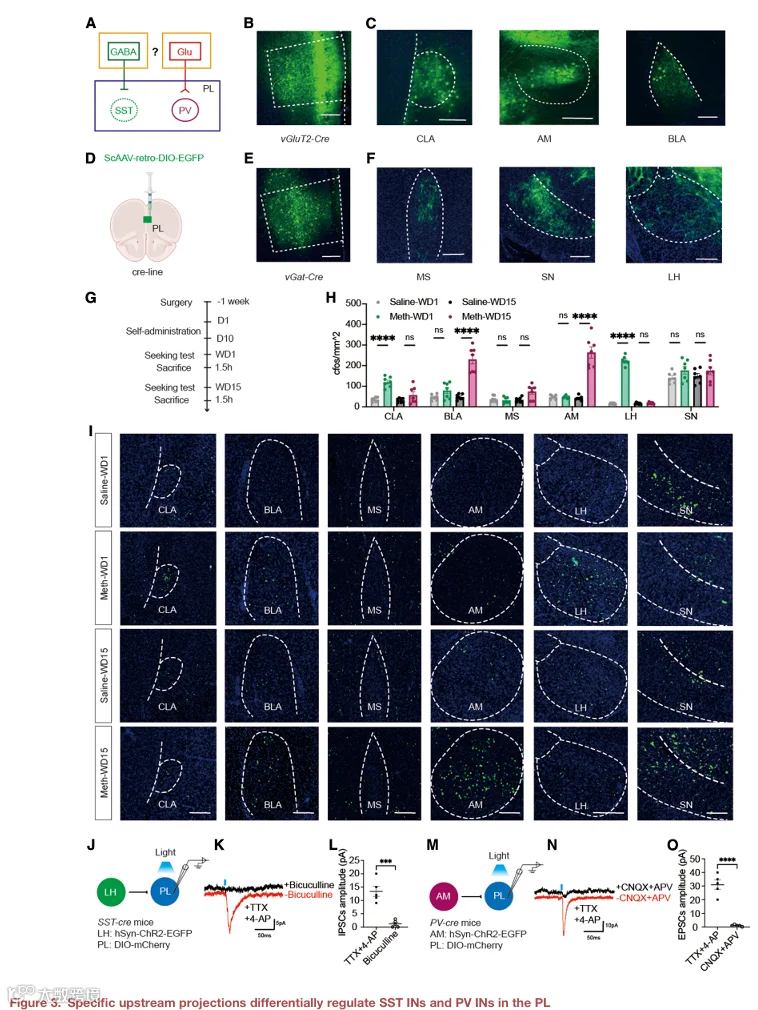
3.特定上游投射介导 PL 中 SST IN 和 PV IN 的差异调节
SST IN 和 PV IN 对谷氨酸能神经元表现出不同的抑制控制:SST IN 降低平均 Ca²⁺ 活动幅度,而 PV IN 在 CNO 注射后降低放电模式(图 S3D–S3G)。这种功能差异可能源于它们不同的传入输入(图 3A)。使用 Cre 依赖的逆行示踪病毒(AAV‑Retro‑DIO‑EGFP)注射到 vGat‑Cre 或 vGluT2‑Cre 小鼠的 PL 中,揭示了 PL 接收的不同 GABA 能(如 LH)和谷氨酸能(如 AM)输入(图 3B、3C、3E 和 3F)。突触素‑mCherry 标记结合 IN 免疫荧光染色显示,LH GABA 能投射优先支配 PL SST IN,而 AM 谷氨酸能投射主要靶向 PV IN(图 S5A–S5L)。
为了识别参与 METH 寻求的上游脑区,我们在 WD1 和 WD15 比较了 METH 与盐水小鼠在线索诱导寻求测试后的全脑 c‑Fos 活动(图 3G)。分析了六个脑区,包括 CLA、BLA、MS、SN、AM 和 LH。两个阶段均显示 BLA 被激活。WD1 还表现出 LH、PVT 和 VP 的活动增加(图 S4M 和 S4N),而 WD15 则显示 AM 的选择性激活(图 3H 和 3I)。考虑到 LH 在奖赏学习中的作用以及 AM 在长时程记忆巩固中的作用,这些发现提示在 METH 寻求过程中存在阶段特异性的环路募集。
使用光遗传学结合全细胞膜片钳记录验证了功能连接。光刺激 LH 投射在 PL SST⁺ 神经元中诱发了短潜伏期的抑制性突触后电流,该电流可被 GABAA 受体拮抗剂荷包牡丹碱以及 TTX 和 4‑AP 阻断,表明其为单突触 GABA 能抑制(图 3J–3L)。类似地,刺激 AM 投射在 PL PV⁺ 神经元中诱发了兴奋性突触后电流,该电流可被 AMPA 受体拮抗剂 CNQX 和 NMDA 受体拮抗剂 APV 以及 TTX 和 4‑AP 阻断,证实了单突触谷氨酸能输入(图 3M–3O)。这些结果确立了 LH→PL SST IN 和 AM→PL PV IN 作为功能上不同的传入环路。
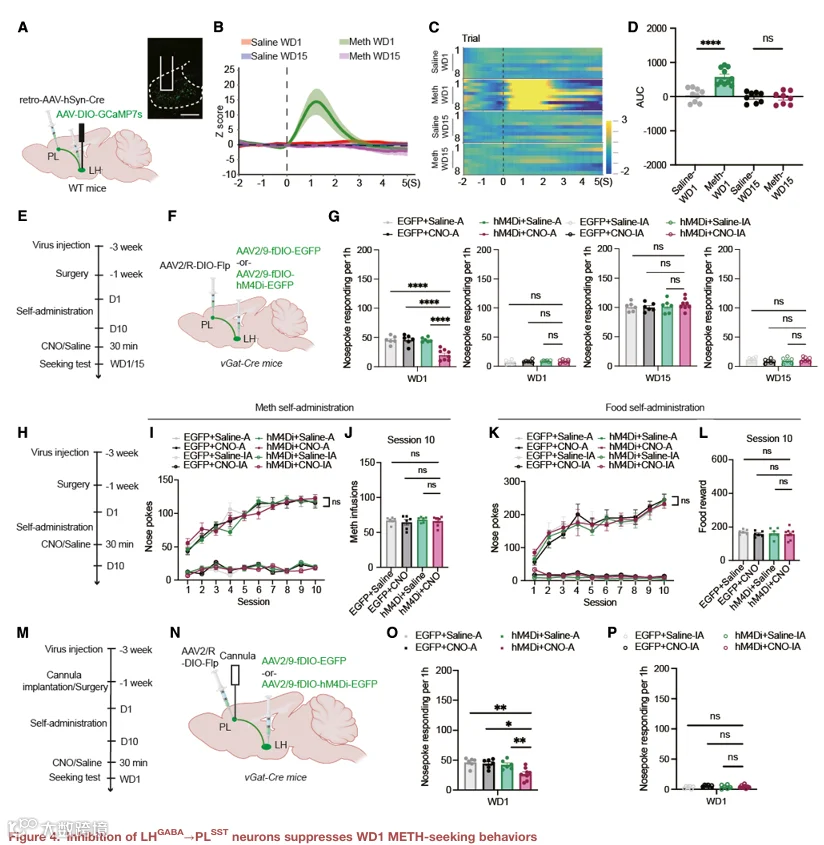
4.抑制 LHᴳᴬᴮᴬ→PL 神经元可抑制 WD1 的 METH 寻求行为
为了直接验证 LH→PL 通路在 WD1 METH 寻求中的必要性,我们在 LH 中注射 Cre 依赖的 hM4Di 病毒,并在 PL 中注射逆行 Cre 病毒,以特异性靶向 LH→PL 投射神经元(图 4A)。光纤光度记录显示,在 WD1 线索诱导寻求测试期间,LH→PL 投射神经元的钙活动显著升高(图 4B–4D)。化学遗传学抑制 LH→PL 投射神经元显著减少了 WD1 的主动鼻触次数,而对照病毒无此效应(图 4E–4G)。该操作对 WD15 的寻求行为无影响(图 4G)。此外,该操作不影响 METH 自身给药的获取(图 4H–4J)或食物自身给药行为(图 4K 和 4L),表明其效应具有行为特异性。
为了验证 LH→PL 通路是否通过 SST IN 发挥作用,我们在 PL 中同时表达 fDIO‑hM4Di 以靶向 SST IN,并在 LH 中表达 Cre 依赖的兴奋性病毒以激活 LH→PL 投射,然后进行光纤光度记录(图 4M–4P)。激活 LH→PL 投射导致 PL SST IN 的钙活动降低(图 4O),而化学遗传学抑制 PL SST IN 则阻断了激活 LH→PL 投射所导致的寻求行为增加(图 4P)。这些数据表明,LH→PL 通路通过抑制 SST IN 来促进 WD1 的 METH 寻求。
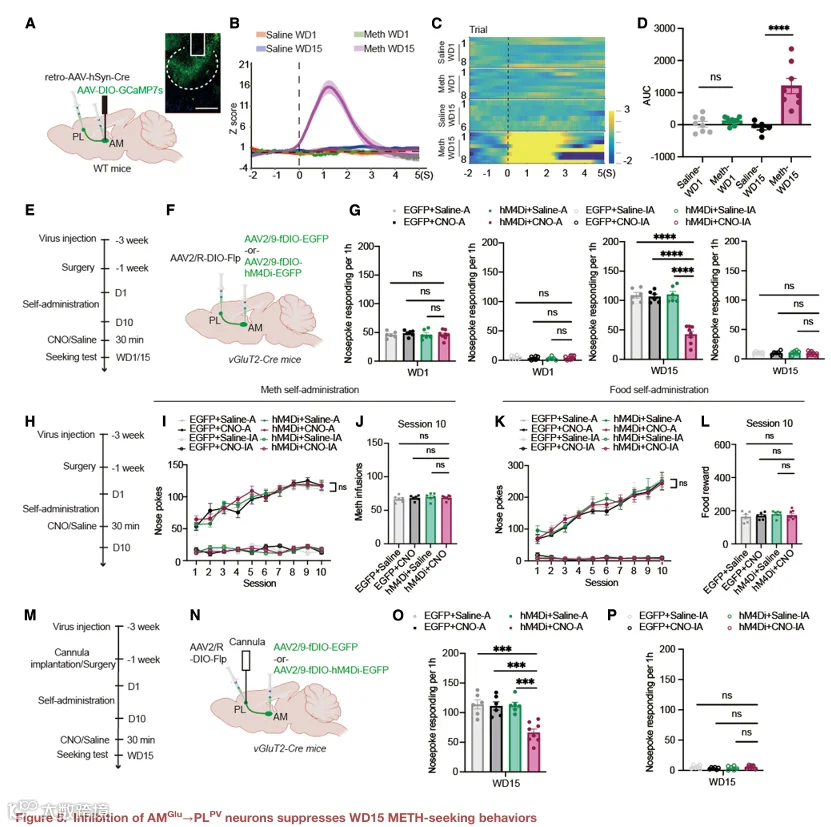
5.抑制 AMᴳˡᵘ→PL 神经元可抑制 WD15 的 METH 寻求行为
接下来,我们研究了 AM→PL 通路在 WD15 METH 寻求中的作用。使用类似的策略特异性靶向 AM→PL 投射神经元(图 5A)。光纤光度记录显示,在 WD15 线索诱导寻求测试期间,AM→PL 投射神经元的钙活动显著升高(图 5B–5D)。化学遗传学抑制 AM→PL 投射神经元显著减少了 WD15 的主动鼻触次数,而对照病毒无此效应(图 5E–5G)。该操作对 WD1 的寻求行为无影响(图 5G)。同样,该操作不影响 METH 自身给药的获取(图 5H–5J)或食物自身给药行为(图 5K 和 5L),表明其效应具有行为特异性。
为了验证 AM→PL 通路是否通过 PV IN 发挥作用,我们在 PL 中同时表达 fDIO‑hM4Di 以靶向 PV IN,并在 AM 中表达 Cre 依赖的兴奋性病毒以激活 AM→PL 投射,然后进行光纤光度记录(图 5M–5P)。激活 AM→PL 投射导致 PL PV IN 的钙活动升高(图 5O),而化学遗传学抑制 PL PV IN 则阻断了激活 AM→PL 投射所导致的寻求行为增加(图 5P)。这些数据表明,AM→PL 通路通过激活 PV IN 来促进 WD15 的 METH 寻求。
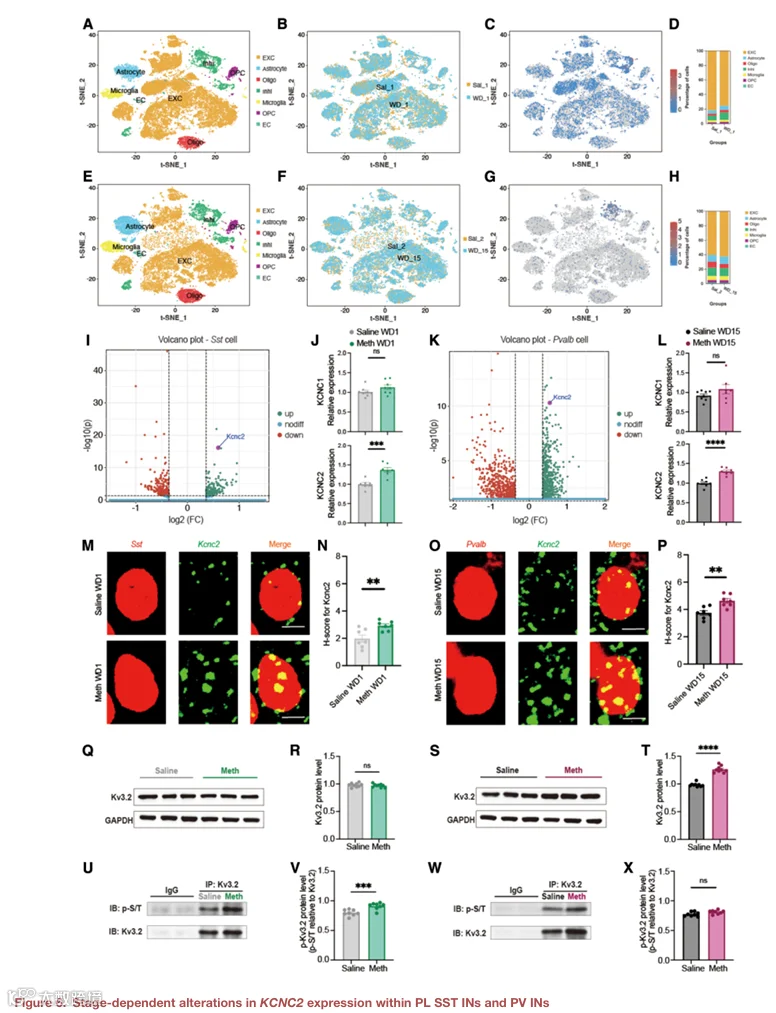
6.PL SST IN 和 PV IN 中 KCNC2 表达的阶段依赖性改变
为了探索 SST IN 和 PV IN 在不同戒断阶段功能差异的分子基础,我们对 WD1 和 WD15 的 METH 与盐水处理小鼠的 PL 进行了单细胞 RNA 测序(图 6A–6H)。t‑SNE 图显示了细胞类型的分布,并分别鉴定了 SST⁺ 和 PV⁺ 细胞群(图 6A–6H)。对 SST⁺ 细胞在 WD1 的差异表达分析显示,与盐水组相比,METH 组中 Kcnc2 是上调最显著的基因之一(图 6I 和 6J)。对 PV⁺ 细胞在 WD15 的分析同样显示 Kcnc2 显著上调(图 6K 和 6L)。定量实时 PCR 验证了 Kcnc2 mRNA 在 WD1 的 SST IN 富集样本和 WD15 的 PV IN 富集样本中均升高(图 S6A–S6D)。此外,免疫荧光染色显示,在 WD1,Kv3.2(由 KCNC2 编码)在 SST IN 中表达升高;在 WD15,Kv3.2 在 PV IN 中表达升高(图 S6E–S6H)。通过免疫共沉淀检测,我们发现 WD1 时 SST IN 中 Kv3.2 的磷酸化水平显著升高,而 WD15 时 PV IN 中 Kv3.2 的磷酸化水平无显著变化(图 S6I 和 S6J)。这些结果表明,KCNC2 在 SST IN 和 PV IN 中以阶段特异性方式被上调,并可能通过不同的翻译后修饰介导其功能。
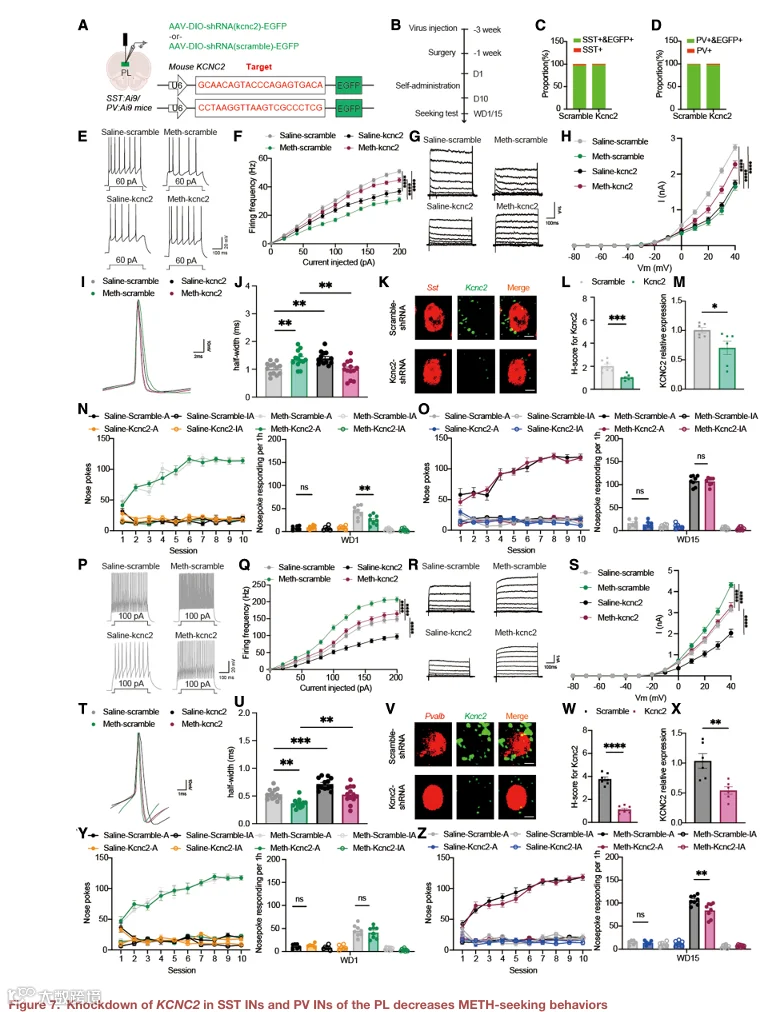
7.在 PL SST IN 和 PV IN 中敲低 KCNC2 可降低 METH 寻求行为
为了直接测试 KCNC2 在 SST IN 和 PV IN 中介导 METH 寻求的功能必要性,我们构建了 Cre 依赖的 shRNA 病毒,在 PL 中特异性敲低 SST IN 或 PV IN 中的 Kcnc2(图 7A)。qPCR 验证了敲低效率(图 S7A 和 S7B)。在 WD1,在 SST IN 中特异性敲低 Kcnc2 显著减少了主动鼻触次数(图 7B 和 7C),而在 PV IN 中敲低 Kcnc2 则无此效应(图 7D)。相反,在 WD15,在 PV IN 中敲低 Kcnc2 显著减少了主动鼻触次数(图 7E 和 7F),而在 SST IN 中敲低 Kcnc2 则无效应(图 7G)。这些效应具有行为特异性,因为敲低不影响 METH 自身给药的获取(图 S7C–S7F)。此外,在 SST IN 中敲低 Kcnc2 挽救了 WD1 时 SST IN 的活性降低,而在 PV IN 中敲低 Kcnc2 挽救了 WD15 时 PV IN 的活性升高(图 S7G–S7J)。这些数据共同表明,KCNC2 在 SST IN 和 PV IN 中以阶段特异性方式调节 METH 寻求行为。
更多结果和补充图表:doi:10.1016/j.neuron.2026.02.017
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!