肥胖青少年2型糖尿病进展迅猛,肾损伤风险极高。代谢手术能否真正“逆转”肾脏病变?最新研究通过单细胞转录组+循环蛋白组学,首次在术前与术后1年的配对肾活检中,揭示了惊人答案:手术显著抑制mTORC1与JAK-STAT信号通路,重编程肾小管代谢,激活AMPK–FOXO3保护轴,同时伴随肾高滤过、肾体积与尿蛋白的显著回落。这项研究不仅为青少年糖肾提供了机制层面的“修复蓝图”,更指明了非手术治疗的新靶点。组学加持,肾脏修复的分子路径首次被清晰描绘。
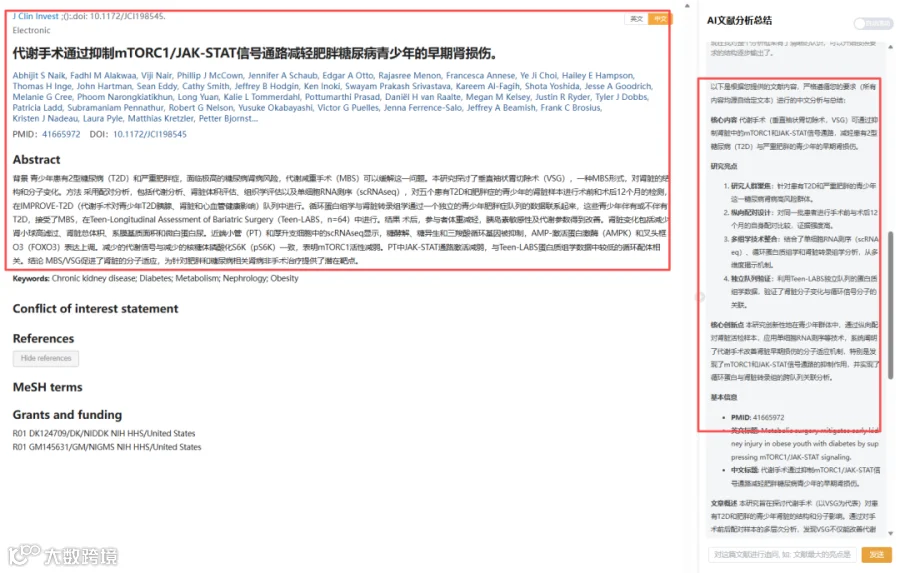
今天给大家解读一篇2月发表在《The Journal of clinical investigation》上的题目为“Metabolic surgery mitigates early kidney injury in obese youth with diabetes by suppressing mTORC1/JAK-STAT signaling.”的文章。本研究在IMPROVE-T2D队列中,对5名患有T2D和肥胖的青少年,在VSG手术前和术后12个月分别采集肾脏活检样本进行配对分析。分析内容包括代谢分析、肾脏体积评估、组织学评估和单细胞RNA测序。同时,将循环蛋白质组学与肾脏转录组学数据,与另一个独立的青少年队列(Teen-LABS,n=64,接受MBS手术,伴或不伴T2D)的数据进行关联分析。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!


团队成员合影(位于上海陆家嘴中心,可随时预约参观)
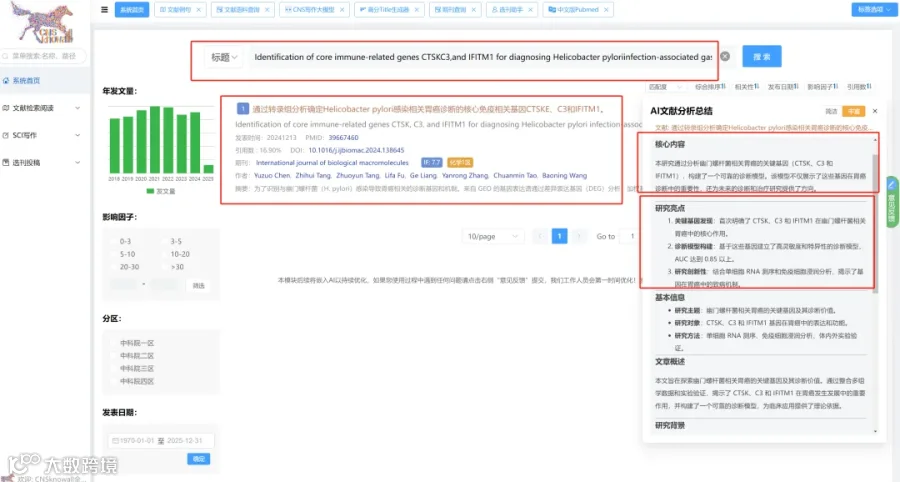
题目:《代谢手术通过抑制mTORC1/JAK-STAT信号通路减轻肥胖糖尿病青少年的早期肾损伤》Metabolic surgery mitigates early kidney injury in obese youth with diabetes by suppressing mTORC1/JAK-STAT signaling
发表期刊:The Journal of clinical investigation
影响因子:13.6
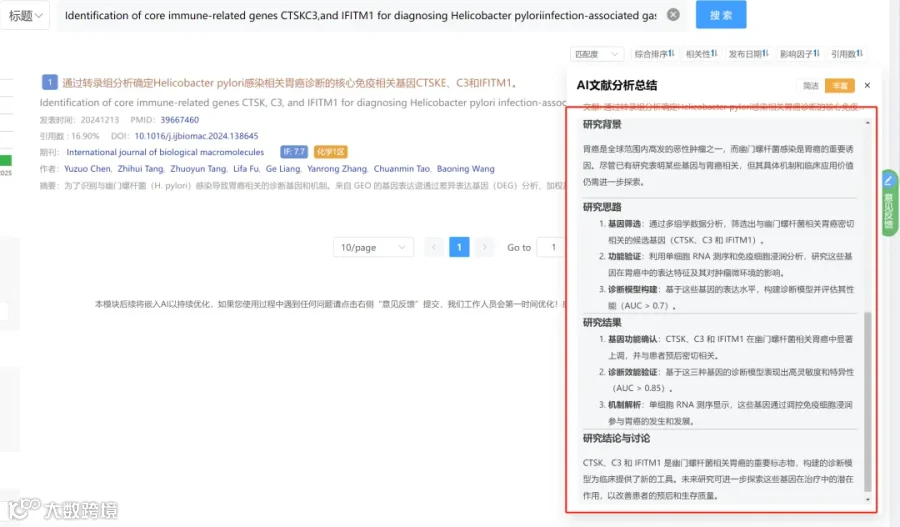
研究背景:
患有2型糖尿病(T2D)和严重肥胖的青少年面临极高的糖尿病肾病风险,而代谢减重手术(MBS)可以缓解这一问题。
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
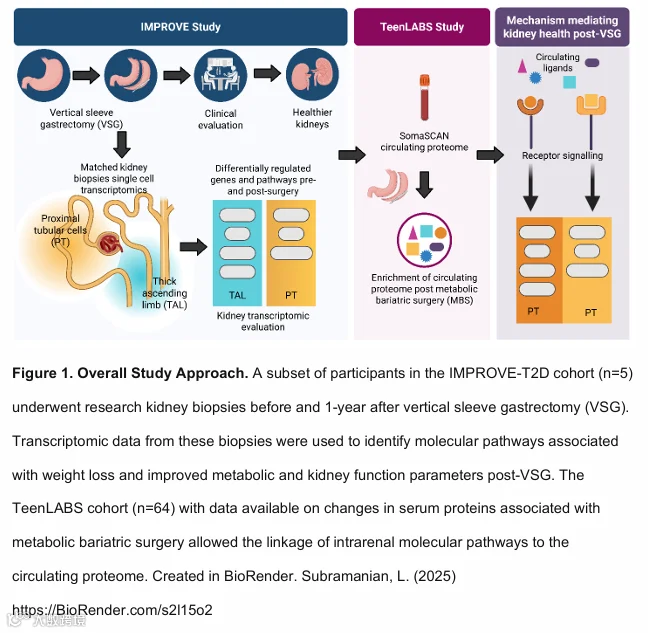
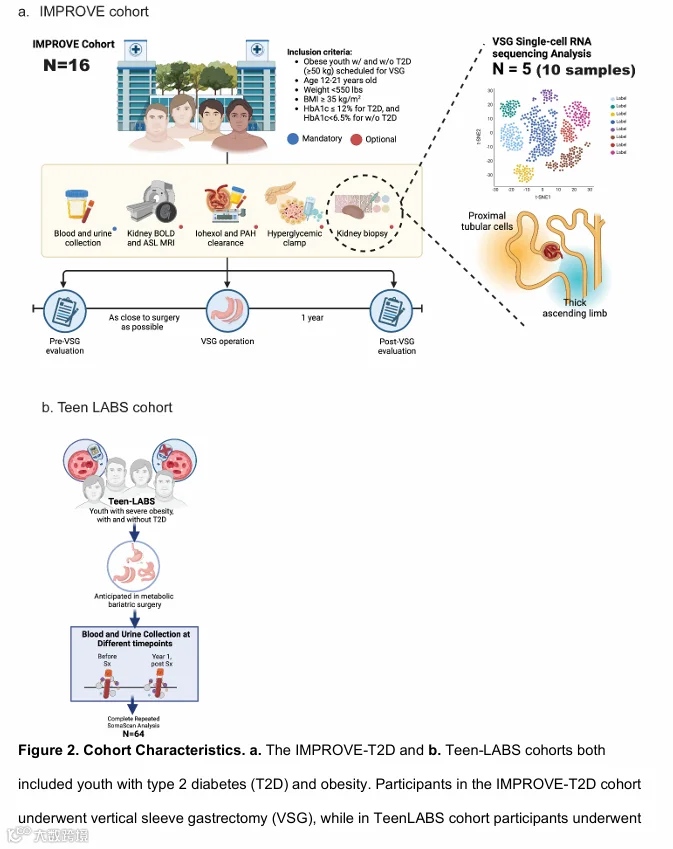
本研究在IMPROVE-T2D队列中,对5名患有T2D和肥胖的青少年,在VSG手术前和术后12个月分别采集肾脏活检样本进行配对分析。分析内容包括代谢分析、肾脏体积评估、组织学评估和单细胞RNA测序。同时,将循环蛋白质组学与肾脏转录组学数据,与另一个独立的青少年队列(Teen-LABS,n=64,接受MBS手术,伴或不伴T2D)的数据进行关联分析。
研究亮点:
- 研究人群聚焦
针对患有T2D和严重肥胖的青少年这一糖尿病肾病高风险群体。
- 纵向配对设计
对同一批患者进行手术前与术后12个月的自身配对比较,证据强度高。
- 多组学技术整合
结合了单细胞RNA测序(scRNAseq)、循环蛋白质组学和肾脏转录组学分析,从多维度揭示机制。
- 独立队列验证
利用Teen-LABS独立队列的蛋白质组学数据,验证了肾脏分子变化与循环信号分子的关联。
研究结果:
- 代谢改善
术后参与者体重减轻,胰岛素敏感性和其他代谢参数得到改善。
- 肾脏结构与功能改善
肾脏变化包括肾小球高滤过减少、肾脏总体积减小、系膜基质面积减少以及微量白蛋白尿减少。
- 分子机制变化
-
近端小管(PT)和厚升支细胞中的scRNAseq显示,糖酵解、糖异生和三羧酸循环相关基因被抑制,而AMPK和FOXO3表达上调。
-
代谢信号减弱与磷酸化S6K(pS6K)减少相一致,提示mTORC1活性减弱。
-
PT细胞中的JAK-STAT通路激活减弱,这与Teen-LABS蛋白质组学数据中较低的循环配体水平相关。
研究总结:
代谢减重手术/垂直袖状胃切除术(MBS/VSG)能够促进肾脏的分子适应,包括抑制mTORC1和JAK-STAT信号通路。这些发现为开发针对肥胖和糖尿病相关肾病的非手术干预措施提供了潜在的分子靶点。
结果译文:
我们在IMPROVE-T2D队列中的五名患有T2D的青少年中,获得了配对的研究性肾活检样本,分别在袖状胃切除术(VSG)前和术后12个月采集(表1)。由于样本量较小,连续变量的数据分布以中位数及其四分位距(IQR,25–75%)表示,未计算p值。BMI下降了31%(中位数从41.5降至28.6 kg/m²),通过碘海醇清除率测得的绝对肾小球滤过率(GFR)下降了23.5%(从228降至175 ml/min),经体表面积(BSA)校正后下降了12.2%(从171降至150 ml/min/1.73m²)。尿白蛋白/肌酐比值(uACR)的中位数下降了57%,而通过MRI评估的总肾脏体积(TKV)下降了13%(从355.9降至319.6 ml)。这些肾脏体积的减少伴随着肾小球核计数(下降20%)、系膜基质面积(下降12.8%)和系膜指数(下降8.4%)的减少。肾小球和系膜体积未见减少。
与此同时,空腹血浆胰岛素水平下降了62.9%(IQR:从17.0降至6.3 mg/dl),糖化血红蛋白(HbA1c)下降了24%(IQR:从6.0降至5.4%),空腹血糖水平下降了18.2%(IQR:从108降至88.3 mg/dl)。胰岛素分泌显著改善,葡萄糖急性胰岛素反应(AIRg)增加了566%(IQR:从49.8升至331.7 μU/mL·min),葡萄糖急性C肽反应增加了350%(IQR:从0.8升至3.6 ng/mL·min)。内源性高胰岛素血症下的葡萄糖处置率(以原始M值衡量)增加了224%(IQR:从2.1升至6.8 mg/kg/min),胰岛素敏感性指数(M/I)增加了60%(IQR:从0.05升至0.08 mg/kg/min per μU/mL)。
这五名患者的临床特征和代谢变化与未提供研究性活检的十一名IMPROVE-T2D参与者相似(补充表S1)。针对VSG后肾细胞中细胞通路评估的具体方法(图1)及各队列概况(图2)已在方法部分进行示意图概述和更详细的描述。
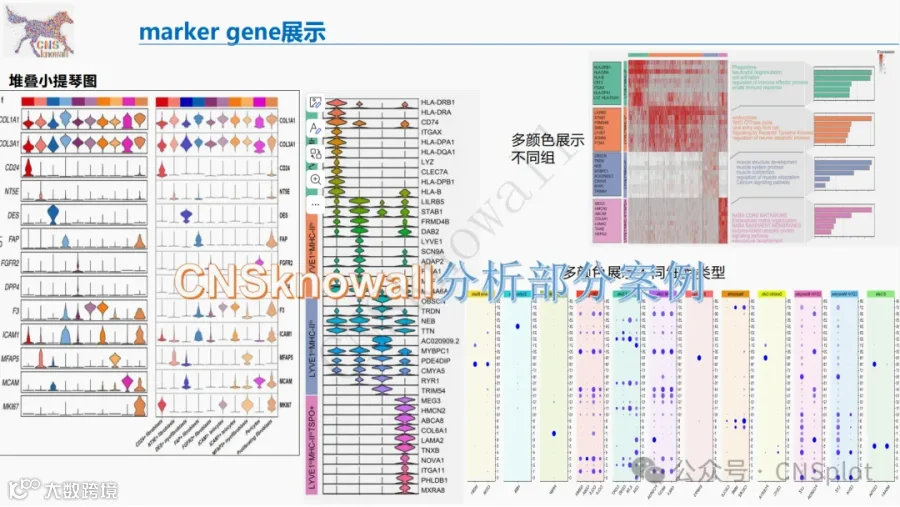
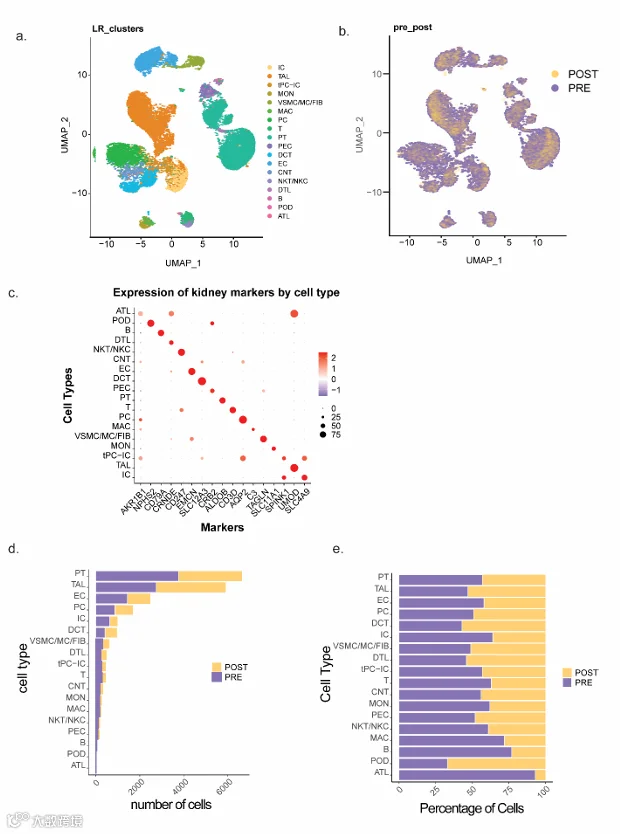
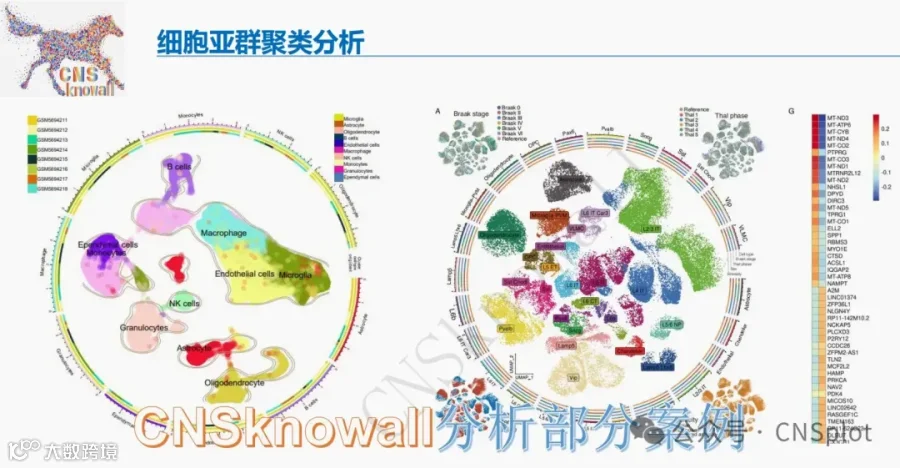
从5对活检样本中,我们分离出23,598个高质量细胞,聚类为18种不同的细胞类型(图3a),术前与术后VSG样本的细胞类型分布相似(图3b)。基于关键细胞类型标志物的表达进行聚类注释(14–16)(图3c)。肾小球细胞类型,如足细胞(18个细胞),因代表性不足而无法进行有意义的分析。不同肾小管上皮细胞以及作为非肾小管内部对照的内皮细胞(EC)的绝对数量(图3d)和百分比(图3e)在术前和术后VSG中相似。补充表S2和S3分别列出了近端小管(PT)和髓袢升支粗段(TAL)细胞中差异表达的基因(DEGs)。
所有比较分析均在术后与术前VSG样本之间进行(n = 23,598个细胞)。健康对照(HC,n = 19,735个细胞,补充图S1)作为参考,用于评估术后VSG样本的变化方向。
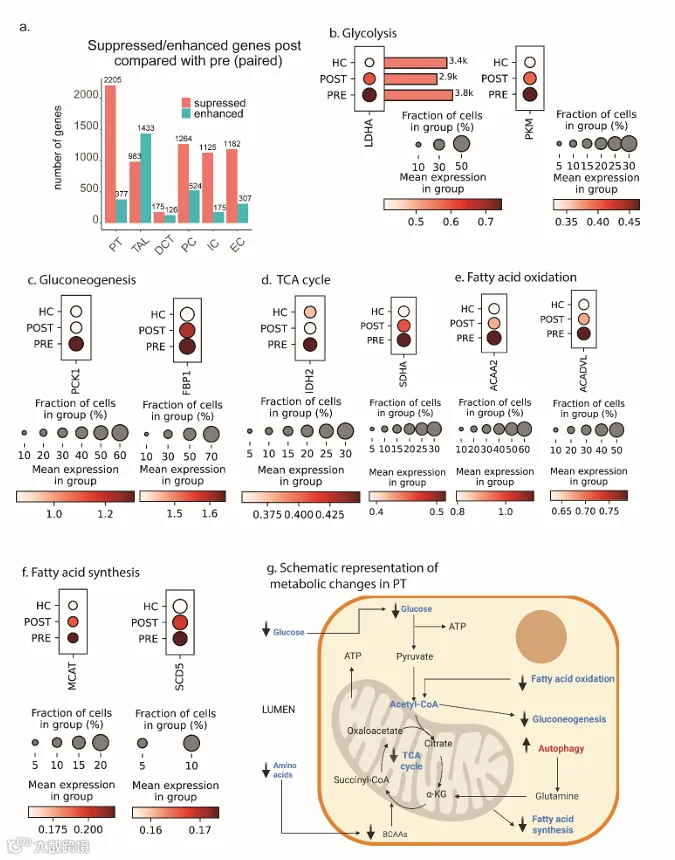
PT和TAL在术前与术后VSG之间的DEGs数量最多(图4a)。PT表现出2582个DEGs,其中术后VSG有2205个基因表达增强,377个基因表达抑制,而TAL有2416个DEGs,其中983个增强,1433个抑制(图4a),这为深入探索这两种肾小管细胞类型提供了强有力的依据。包括远曲小管(DCT)、主细胞(PC)、闰细胞(IC)和内皮细胞在内的远端肾单位节段也显示出表达差异。使用Reactome对PT中下调基因进行的通路富集分析(补充图S2a)发现,多个代谢相关通路减少,包括三羧酸循环、氧化磷酸化、糖酵解和糖异生。与蛋白质翻译和细胞应激反应相关的通路也减少。为了在基因水平上展示这一点,评估了代谢相关通路代表性基因的表达变化。术后VSG后,参与糖酵解的基因表达降低,如乳酸脱氢酶A(LDHA,p < 0.0001)、丙酮酸激酶(PKM,p < 0.0001)(图4b);参与糖异生的基因如磷酸烯醇式丙酮酸羧激酶1(PCK1,p < 0.0001)、果糖-1,6-二磷酸酶1(FBP1,p = 0.01)(图4c);参与三羧酸循环的基因如异柠檬酸脱氢酶2(IDH2,p < 0.0001)、琥珀酸脱氢酶A(SDHA,p < 0.0001)(图4d);参与脂肪酸氧化的基因如乙酰辅酶A酰基转移酶2(ACAA2,p < 0.0001)、极长链酰基辅酶A脱氢酶(ACADVL,p < 0.0001)(图4e)。参与脂肪酸合成的线粒体丙二酰辅酶A酰基载体蛋白转酰基酶(MCAT,p = 0.008)和硬脂酰辅酶A去饱和酶5(SCD5,p < 0.0001)的表达也出现下降(图4f)。图4g示意性总结了PT中观察到的转录变化,包括术后VSG后雷帕霉素机制靶点复合物1(mTORC1)信号传导的减少(后续详述)。在PT中增强的基因中,发现与发育生物学和受体酪氨酸激酶(RTK)信号传导相关的通路增加(补充图S2b)。
与PT类似,TAL细胞在VSG后也表现出代谢相关转录本的下调。对TAL中983个抑制基因(图4a)的通路富集分析发现,三羧酸循环、呼吸电子传递、通过化学渗透偶联合成ATP、通过解偶联蛋白产热是最富集的通路,同时蛋白质翻译相关通路也下调(补充图S3a)。
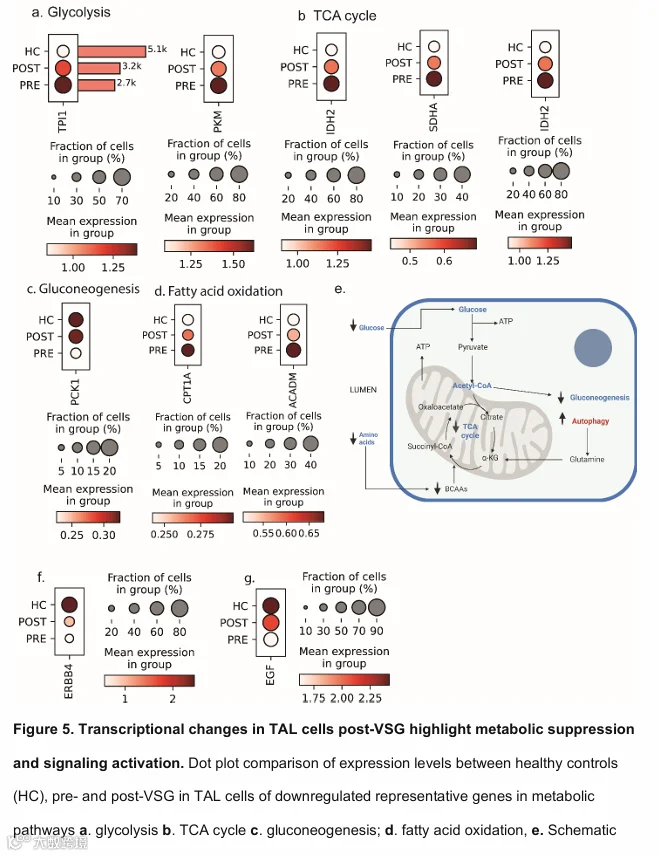
与PT细胞一样,TAL细胞中富集代谢通路的代表性基因显示,与糖酵解相关的基因表达降低(图5a),包括磷酸丙糖异构酶(TPI1,p ≤ 0.0001)和PKM(p < 0.0001)。此外,与三羧酸循环相关的基因如IDH2(p < 0.0001)和SDHA(p = 0.003)(图5b);与糖异生相关的基因如PCK1(p = 0.002,图5c);与脂肪酸氧化相关的基因如肉碱棕榈酰转移酶1(CPT1A,p < 0.0001)和中链酰基辅酶A脱氢酶(ACADM,p < 0.0001)均被抑制(图5d)。图5e的示意图总结了VSG后TAL中的转录变化。TAL细胞中显著上调的基因包括Erb-B2受体酪氨酸激酶4(ERBB4,图5f)和表皮生长因子(EGF,图5g),这与促进肾小管恢复的信号传导一致。与PT细胞一样,受体酪氨酸激酶信号传导和信号转导是上调最显著的通路之一(补充图S3b)。
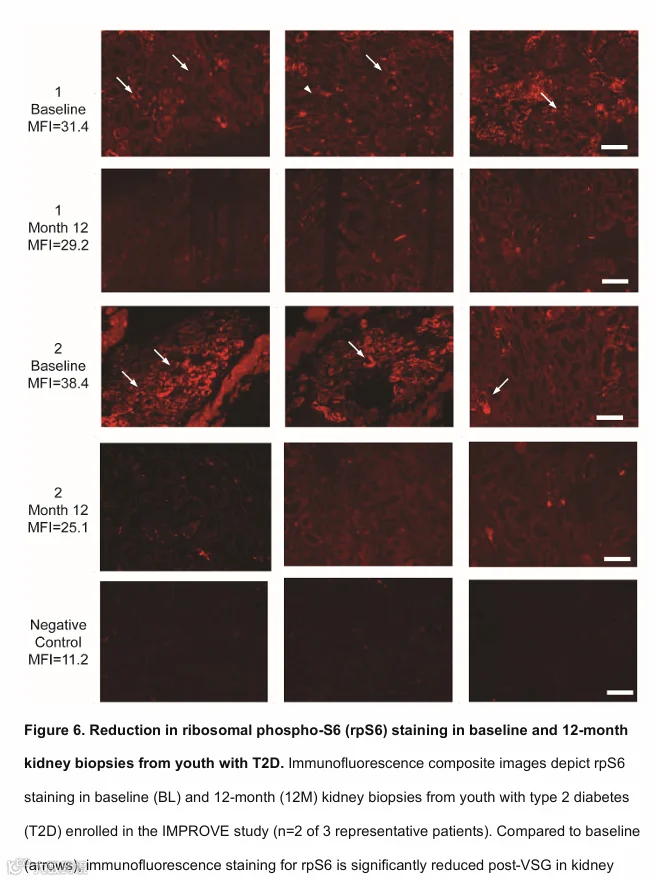
基于转录组变化和观察到的生理反应强烈提示VSG后PT和TAL中mTORC1信号传导减少,我们通过免疫荧光(IF)染色评估了肾组织中mTORC1信号通路活性,染色目标为mTORC1-S6K1通路的下游底物——核糖体磷酸化S6(rpS6 S240/S244)(图6,两名代表性患者)。在术前VSG样本中,三名研究患者的14个切片中,平均荧光强度(MFI,任意单位)为34.9。免疫荧光染色主要定位于肾小管上皮细胞和肾小球,包括足细胞,与mTORC1通路活性一致。相比之下,术后12个月,来自这三名患者的15个切片中rpS6 IF强度较低,为28.3(p = 0.19),且空间分布减少。
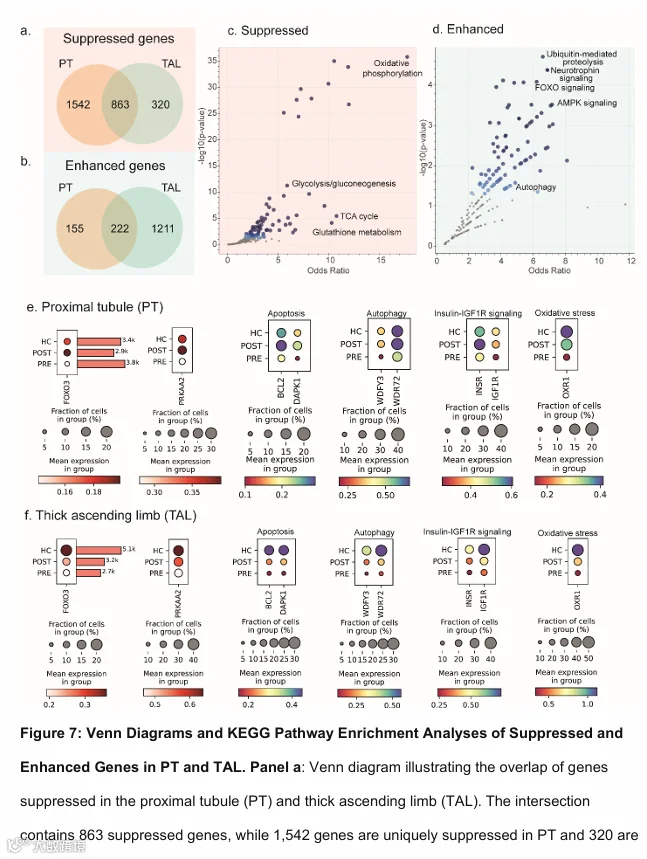
鉴于PT和TAL在VSG后基因表达和通路变化的相似性,特别是在代谢和信号通路方面(补充图S2和S3),我们检查了这两种细胞类型之间DEGs的重叠情况,以识别跨肾单位的核心作用机制。在PT和TAL中,VSG后共有863个基因下调,222个基因上调(图7a和b)。使用京都基因与基因组百科全书(KEGG,2021版)(17)对这些DEGs进行通路富集分析。排名靠前的下调通路包括氧化磷酸化(OR: 17.08,p ≤ 0.0001)、三羧酸循环(OR: 10.72,p < 0.0001)、糖酵解/糖异生(OR: 5.81,p = 0.0001)和谷胱甘肽信号通路(OR: 4.12,p = 0.02)(补充表S4,图7c)。排名靠前的上调通路包括泛素介导的蛋白水解(OR: 6.34,p = 0.004)、神经营养因子信号通路(OR: 6.62,p = 0.004)、叉头盒蛋白O3(FOXO3)介导的信号通路(OR: 5.97,p = 0.004)和腺苷单磷酸活化蛋白激酶(AMPK)信号通路(OR: 5.67,p = 0.007)(补充表S4,图7d)。
在重叠分析中,AMPK的催化亚基α2(PRKAA2)和FOXO3在PT和TAL中均上调(图7e和7f,补充表S5)。然后我们进一步评估了已知受AMPK-FOXO3调控的过程(18)的表达。除了前面讨论的与代谢适应相关的基因外,VSG后,与凋亡相关的基因如BCL2凋亡调节因子(BCL2,p < 0.05)、死亡相关蛋白激酶1(DAPK1,p < 0.05);自噬相关的基因如含WD重复和FYVE结构域蛋白3(WDFY3,p < 0.05)、WD重复结构域蛋白72(WDR72,p < 0.05);胰岛素受体信号(INSR,p < 0.05)、胰岛素样生长因子1受体(IGF-1R,p < 0.05);以及抗氧化应激相关的基因如抗氧化抵抗蛋白1(OXR1,p < 0.05)的表达均上调。与这些数据一致,关键细胞过程如自噬(OR: 4.17,p = 0.02)、凋亡(OR: 4.01,p = 0.03)、胰岛素信号通路(OR: 4.91,p = 0.01)和长寿调控通路(与IGF-1R信号密切相关,OR: 5.7,p = 0.01)均上调(补充表S6)。线粒体自噬也呈增加趋势(OR: 4.25,p = 0.10)。
为了支持肾小管细胞中观察到的代谢重编程反映了VSG后血清葡萄糖、氨基酸和生长因子减少的假说(19–24),我们进行了体外实验,检测了上皮基础培养基系列稀释对HK-2人肾上皮细胞的影响。进行性营养剥夺诱导了pS6K1/S6K1比值的逐步降低和相应的pAMPK/AMPK比值的增加(补充图S4)。在2.5%培养基浓度下,mTORC1活性几乎被消除,与50 ng/mL雷帕霉素相当。重要的是,pAkt/Akt比值保持不变(约0.83–0.98),表明mTORC1活性降低反映了直接的营养感应,而非Akt/PI3K介导的效应。严重的营养剥夺也诱导了强烈的AMPK活化(与雷帕霉素组的4倍相比,达到10–12倍)。这些体外发现重现了我们的体内观察结果,并为VSG后热量摄入减少直接调节肾小管上皮细胞中mTORC1和AMPK信号传导提供了机制支持。
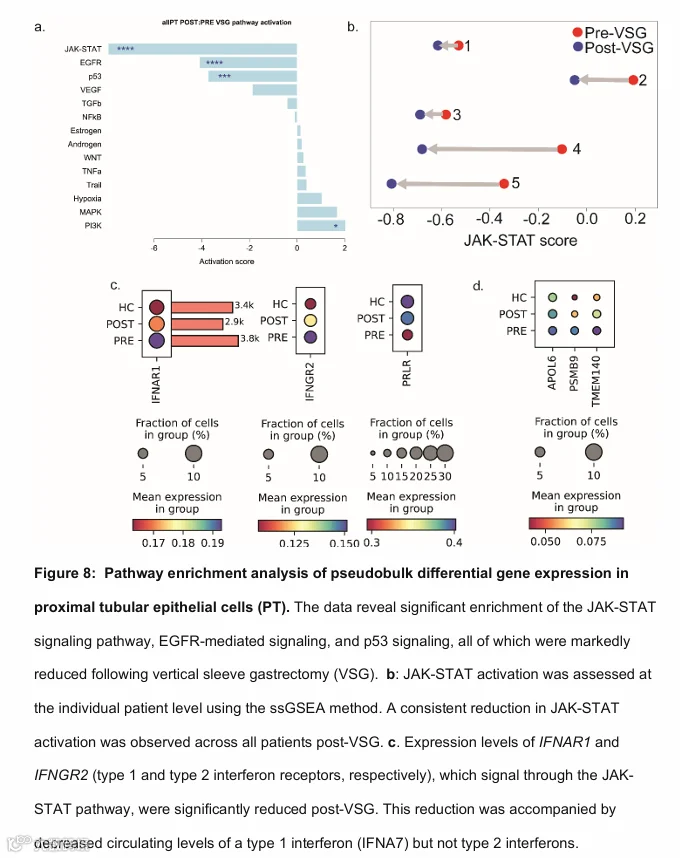
为了评估VSG后受影响的其他通路,重点关注具有治疗意义的信号通路,我们使用了通路响应基因(PROGENy)工具。对PT伪批量数据的PROGENy分析显示,VSG后失调的主要信号通路是Janus激酶/信号转导和转录激活通路(JAK-STAT)、表皮生长因子受体(EGFR)和p53信号通路(图8a)。先前发表的量化下游JAK-STAT通路活性的17基因JAK-STAT鼻内特征(25, 26)显示,每位参与者的JAK-STAT活性评分从术前到术后VSG均有所下降(图8b,补充表S7)。
为了评估观察到的肾内JAK-STAT信号减少是否与VSG后全身循环中JAK-STAT配体的减少相关,我们分析了一个更大规模队列(Teen-LABS;n = 64)的数据,该队列在MBS前两周内和术后一年收集了血浆SOMAscan蛋白质组数据(13)。补充表S8总结了该Teen-LABS队列的人口统计学和临床信息。
表2总结了Teen-LABS队列中VSG后通过JAK-STAT信号传导的循环配体水平的变化,这些变化达到了假发现率(FDR)阈值< 0.05。循环水平显著降低的配体包括细胞因子,如白细胞介素-2(IL2)、IL6、IL23和干扰素α-7(IFNA7)(27–30)。此外,我们观察到循环生长激素受体(GHR)水平的降低(GHR是SOMAscan平台上生长激素生物活性的标志(31))以及催乳素(PRL)水平的降低,两者结合的受体均通过JAK-STAT信号传导(32–34)。
在评估PT中这些循环配体的相应受体时,我们观察到I型干扰素受体(IFNAR1,p < 0.0001)和II型干扰素受体(IFNGR2,p < 0.0001)的表达降低(图8c),这与来自全身来源而非PT内产生的配体的活性信号减少一致。其他循环配体(包括IL2、IL6、IL12、IL23和瘦素)的已知JAK-STAT信号受体转录本在PT中未表达。GHR蛋白在PT中表达(人类蛋白质图谱),但GHR mRNA在PT中无差异表达。然而,VSG后PT中催乳素受体(PRLR)的表达显著增加(图8c)。最后,图8d展示了17基因JAK-STAT肾内特征中3个代表性基因的表达变化,作为图8b中评分的潜在变化示例。
更多结果和补充图表:doi:10.1172/JCI198545
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!