本研究利用GeoMx数字空间多组学技术(DSP),对涵盖食管鳞癌(ESCC)从正常上皮、癌前病变到转移淋巴结全过程的32例组织样本进行了深入分析。研究揭示了肿瘤启动阶段表皮发育与角化分化的协同失调,以及晚期PI3K/AKT通路的持续激活与三级淋巴结构(TLS)功能障碍。通过生信分析与实验验证,锁定仅在晚期高表达且预示不良预后的关键驱动基因——OGT。该基因通过增强O-GlcNAc糖基化修饰,显著提升ESCC细胞的迁移侵袭能力。本文为理解ESCC演化过程中的肿瘤生态系统动态提供了精准治疗新靶点。
今天给大家解读一篇3月发表在《Cell Reports Medicine》上的题目为“Spatial omics study reveals molecular-cellular dynamics of tumor ecosystem in esophageal squamous-cell carcinoma initiation and progression.”的文章。本研究对18名患者处于不同ESCC阶段的32个组织样本进行了数字空间分析。研究发现,在ESCC起始阶段,上皮细胞发育和角质细胞分化途径发生协调性调控,伴随B细胞增加和T细胞减少。在晚期阶段,PI3K/AKT信号通路持续上调,三級淋巴结构功能失调。O-GlcNAc转移酶基因仅在晚期被特异性激活,其表达与不良预后相关,敲低该基因可抑制ESCC细胞的迁移和侵袭。该研究通过空间组学揭示了ESCC生态系统演化机制,为精准治疗提供了路线图。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!包括数据集、绘图代码、图表复现、思路总结、参考文献……0代码!鼠标点点点即可轻松完成5-10分生信SCI全文复现!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!



团队成员合影(位于上海陆家嘴中心,可随时预约参观)
题目:《空间组学研究揭示了食管鳞状细胞癌发生和发展过程中肿瘤生态系统的分子-细胞动态》Spatial omics study reveals molecular-cellular dynamics of tumor ecosystem in esophageal squamous-cell carcinoma initiation and progression
发表期刊:Cell Reports Medicine
影响因子:10.6
研究背景:
食管鳞状细胞癌是东亚地区主要的食管癌亚型,具有进展快、预后差的特点。既往研究多关注ESCC起始阶段的分子指标,或主要聚焦于肿瘤细胞本身,常忽略肿瘤微环境。从正常组织到食管鳞状癌前病变,最终发展为肿瘤和转移的整个进化过程中的细胞和分子机制尚不清楚。传统的批量或单细胞RNA测序技术无法保留空间信息。本研究旨在利用数字空间分析技术,系统表征ESCC发生发展全过程中上皮细胞和免疫微环境的分子改变。
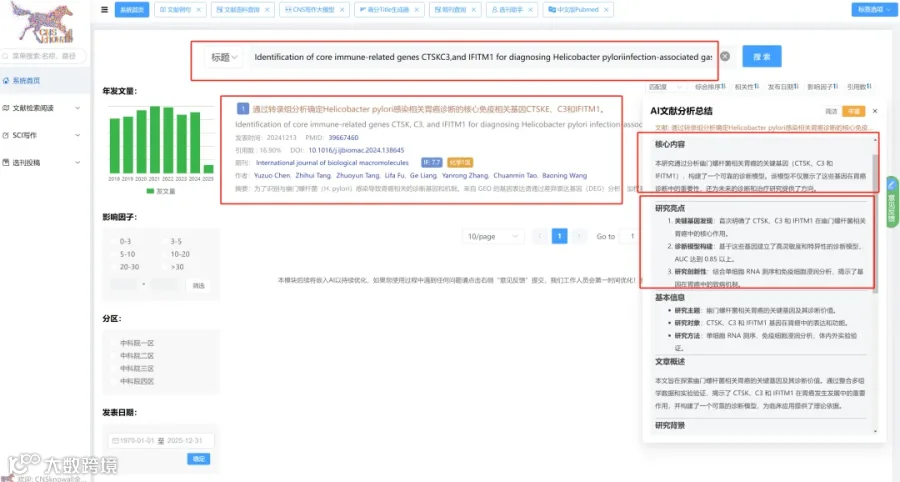
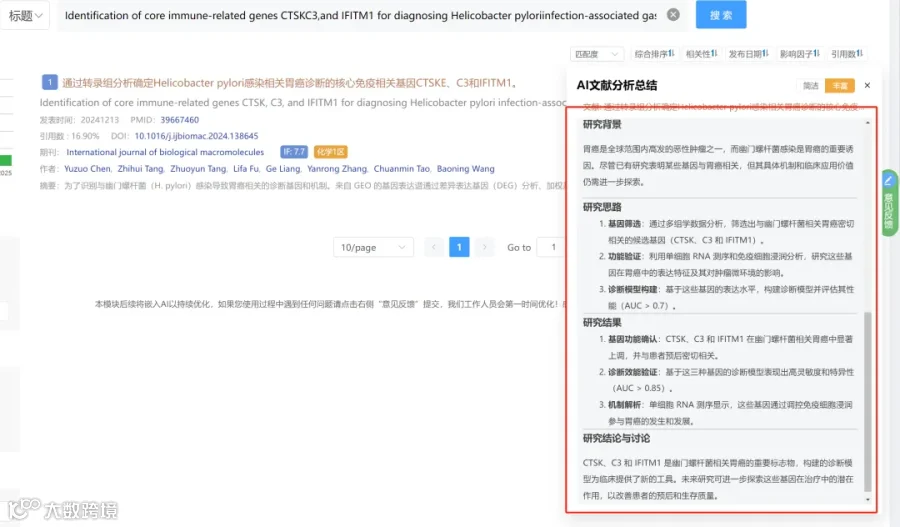
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
- 样本与分组
前瞻性收集18名患者不同ESCC阶段的32个组织样本,包括正常上皮、食管鳞状癌前病变、非转移性ESCC、转移性ESCC和转移淋巴结组织。
- 空间转录组分析
使用基于数字空间分析技术的空间全转录组图谱面板,针对每个感兴趣区域,划分并分别定量分析了上皮细胞富集区、巨噬细胞富集区、中性粒细胞富集区、基质富集区和淋巴结构区。
- 多层次分析
分别分析了上皮区室在肿瘤起始和进展中的基因表达与通路变化,以及肿瘤微环境中免疫细胞组成、三級淋巴结构状态和细胞间通讯的动态演变。
- 关键基因功能验证
通过免疫组化、多重免疫荧光、细胞功能实验(增殖、迁移、侵袭)和体内外模型(皮下移植瘤、肺转移模型),验证了O-GlcNAc转移酶在ESCC进展中的促癌功能。
- 机制探索
结合RNA测序、蛋白质组学和糖蛋白质组学分析,探索O-GlcNAc转移酶的作用机制。
研究亮点:
- 涵盖全进程的空间分析
对从ESCC起始到进展的32个组织样本进行了数字空间分析。
- 明确早期特征
- 揭示进展驱动力
发现PI3K/AKT信号通路的持续激活和三級淋巴结构功能障碍共同促进ESCC演进。
- 发现关键预后因子
O-GlcNAc转移酶在晚期特异性激活,驱动肿瘤进展并与不良预后相关。
研究结果:
ESCC起始阶段的上皮细胞动态:
-
从正常上皮到癌前病变,表皮发育途径激活,角质细胞分化途径被抑制,伴随胚胎样基因(如SOX4, KRTDAP)上调。
-
从癌前病变到早期癌症,细胞外基质-受体相互作用和PI3K/AKT信号通路持续激活,同时天然免疫和B细胞介导的免疫反应相关基因在癌前病变中高表达而在早期癌症中下调。
ESCC起始阶段的肿瘤微环境变化:
- B细胞和浆细胞
- CD4+和CD8+ T细胞
-
在三級淋巴结构中,APOBEC3A基因表达随着病变进展显著增加,且与B细胞激活标志物IGHM正相关。
ESCC进展到晚期的特征:
-
上皮区室中,PI3K/AKT信号通路、细胞迁移/黏附/增殖相关通路持续上调;营养反应相关基因(如OGT, TFRC)仅在晚期激活。
- O-GlcNAc转移酶
被鉴定为晚期特异性基因,其高表达与ESCC患者不良预后显著相关。功能实验证实,OGT敲低抑制ESCC细胞增殖、迁移和侵袭,而过表达则增强这些恶性表型。体内实验显示OGT敲低抑制肿瘤生长,过表达促进肺转移。
ESCC进展中肿瘤微环境的重塑:
-
晚期阶段,肥大细胞、中性粒细胞和内皮细胞比例大幅下降,巨噬细胞有增加趋势。
-
细胞间通讯分析发现,基质区室通过ACTIVIN、CHEMERIN和PERIOSTIN通路与上皮/免疫区室的相互作用在晚期特异性增强或信号强度增加。
-
三級淋巴结构在晚期出现功能障碍,表现为免疫细胞招募相关的细胞因子(如CCL21)表达受抑制,而与肿瘤转移相关的基因(如TNC)表达上调。
OGT的作用机制:
-
OGT高表达组富集于细胞周期、DNA复制、JAK-STAT和Wnt等促肿瘤通路。
-
-
OGT过表达通过O-GlcNAc糖基化修饰增强AKT Ser473位点的磷酸化。
研究总结:
本研究系统揭示了ESCC发生发展过程中,上皮细胞与肿瘤微环境各组分协同演化的时空动态。核心结论包括:
- 起始机制
表皮发育和角质细胞分化的协调性失调是ESCC起始的关键,伴随以B细胞增多和T细胞减少为特征的早期免疫微环境改变。
- 进展驱动力
PI3K/AKT信号通路的持续激活是贯穿ESCC演进的核心驱动力。
- 关键分子
O-GlcNAc转移酶是ESCC晚期特异性激活的关键分子,通过糖基化修饰驱动肿瘤进展,是潜在的预后标志物和治疗靶点。
- 微环境演变
肿瘤微环境经历动态重塑,晚期出现三級淋巴结构功能障碍和以ACTIVIN、CHEMERIN、PERIOSTIN通路介导的基质-上皮/免疫细胞通讯增强。
局限性:
-
所有癌前病变均为高级别上皮内瘤变,未纳入低级别病变患者,限制了向超早期风险分层或生物标志物开发的直接转化。
-
多重免疫荧光的验证队列与数字空间分析为同一患者队列,缺乏独立的外部验证队列,可能影响发现的普适性。
结果译文:
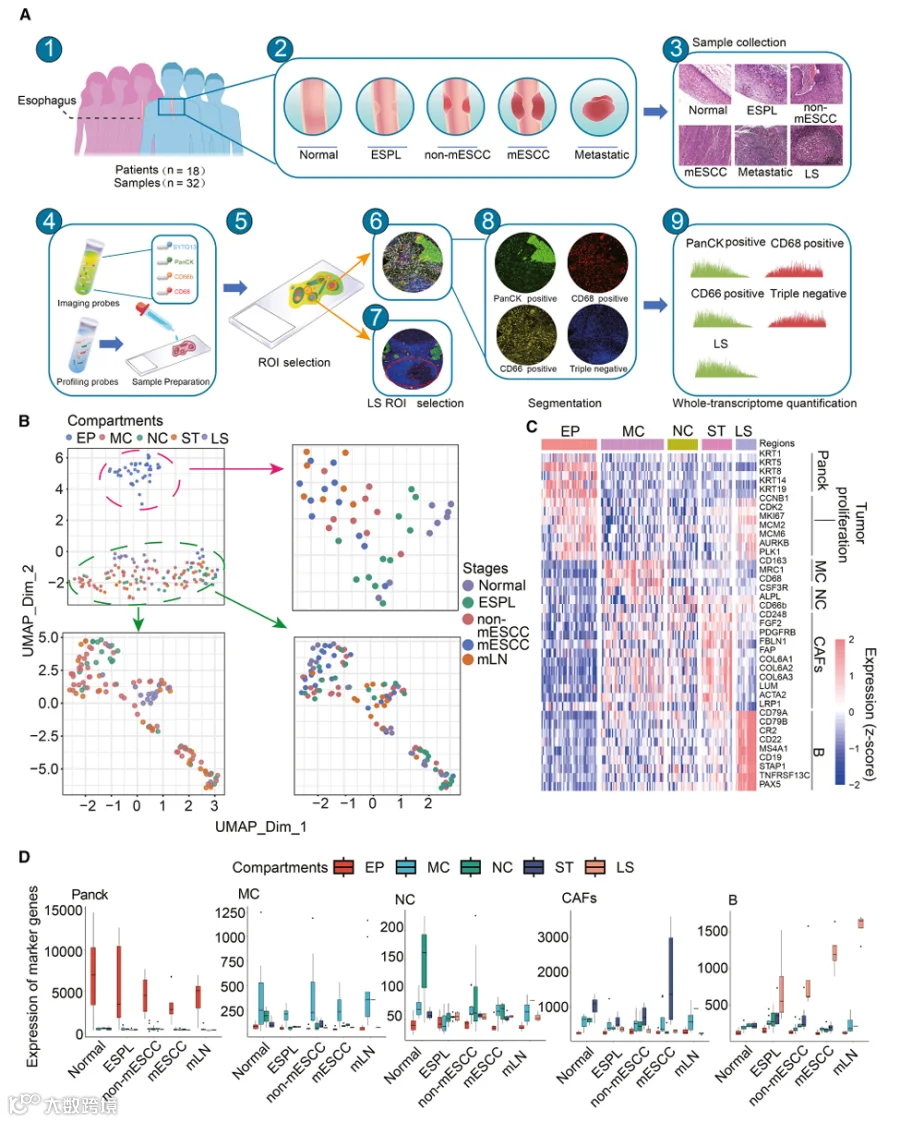
我们前瞻性地收集了18名处于ESCC起始和进展不同阶段患者的32份组织样本,包括正常上皮(NE)、食管鳞状上皮癌前病变(ESPL)、非转移性ESCC(non-mESCC)、转移性ESCC(mESCC)和转移性淋巴结(mLN)组织(表1)。为了系统地表征ESCC演化过程中上皮细胞和免疫微环境的分子改变,我们使用了基于DSP技术的空间全转录组图谱(WTA)panel。如图1A所示,DSP包括免疫荧光抗体探针杂交、感兴趣区域(ROI)选择和分割,以及通过测序进行全转录组定量。为了专门研究各组分的分子特征,我们聚焦于上皮和间质,并对每个ROI内的上皮细胞和免疫细胞区室进行了分割。此外,根据苏木精和伊红(H&E)染色,还选择了具有淋巴结构(LSs;包括肿瘤组织中的TLSs和mLN中的淋巴滤泡)的ROI。在每个ESCC阶段,识别出五种类型的感兴趣区域(AOs)并分别使用全转录组测序进行定量,包括上皮细胞富集(EP,PanCK阳性)、巨噬细胞富集(MC,CD68阳性)、中性粒细胞富集(NC,CD66b阳性)、间质富集(ST,三阴性)和LS AOs。经过严格的技术信号和背景质量控制,共保留165个AO用于后续分析(表S1和S2)。
基于所有空间定义的AOs进行均匀流形近似和投影(UMAP)降维分析(图1B)。正如预期,观察到EP区室和间质微环境区室之间的清晰分离。使用UMAP对EP AOs的进一步亚聚类分析揭示了所分析区室与ESPL和ESCC进展阶段的组织学特征之间的密切关联。此外,我们观察到non-mESCC EP区室位于ESPL和mESCC或mLNs的EP区室之间。同时,mESCC和mLNs聚集在一起,表明它们共享相似的转录程序谱。相反,在来源于间质微环境区室的不同分割区室之间未观察到明显区分,表明间质细胞在ESPL和ESCC发展中可能发挥更复杂的调控作用。然后,我们基于经典标志物进一步评估了五种类型AOs中的基因表达程序(图1C)。与我们的分割策略一致,五个分割组织区域的基因表达与其代表性细胞类型基本对应。相应地,PanCK、巨噬细胞、中性粒细胞、癌症相关成纤维细胞和B细胞基因程序的高表达模式分别在EP、MC、NC、ST和LS区室中观察到(图1D)。此外,我们采用xCell来量化不同区室中上皮细胞、间质细胞、巨噬细胞和中性粒细胞的富集分数。每个区室主要细胞类型的富集分数始终与该区室的预定义身份一致(图S1A)。此外,我们使用CIBERSORT对我们DSP数据的非上皮区域进行了解卷积分析,并利用scRNA-seq衍生的LM22特征矩阵来量化巨噬细胞和中性粒细胞的比例。结果显示,与其他区室相比,巨噬细胞富集区室中的巨噬细胞比例显著更高,而中性粒细胞富集区室中的中性粒细胞比例显著升高(图S1B)。这些结果表明,基于DSP技术,来自不同组织的区域被准确地分割和量化,这使我们能够在后续研究中系统且准确地分析ESCC起始和进展过程中的分子变化。
2.空间转录组表达模式揭示EP区室中肿瘤发生通路的激活与ESCC的逐步起始相关
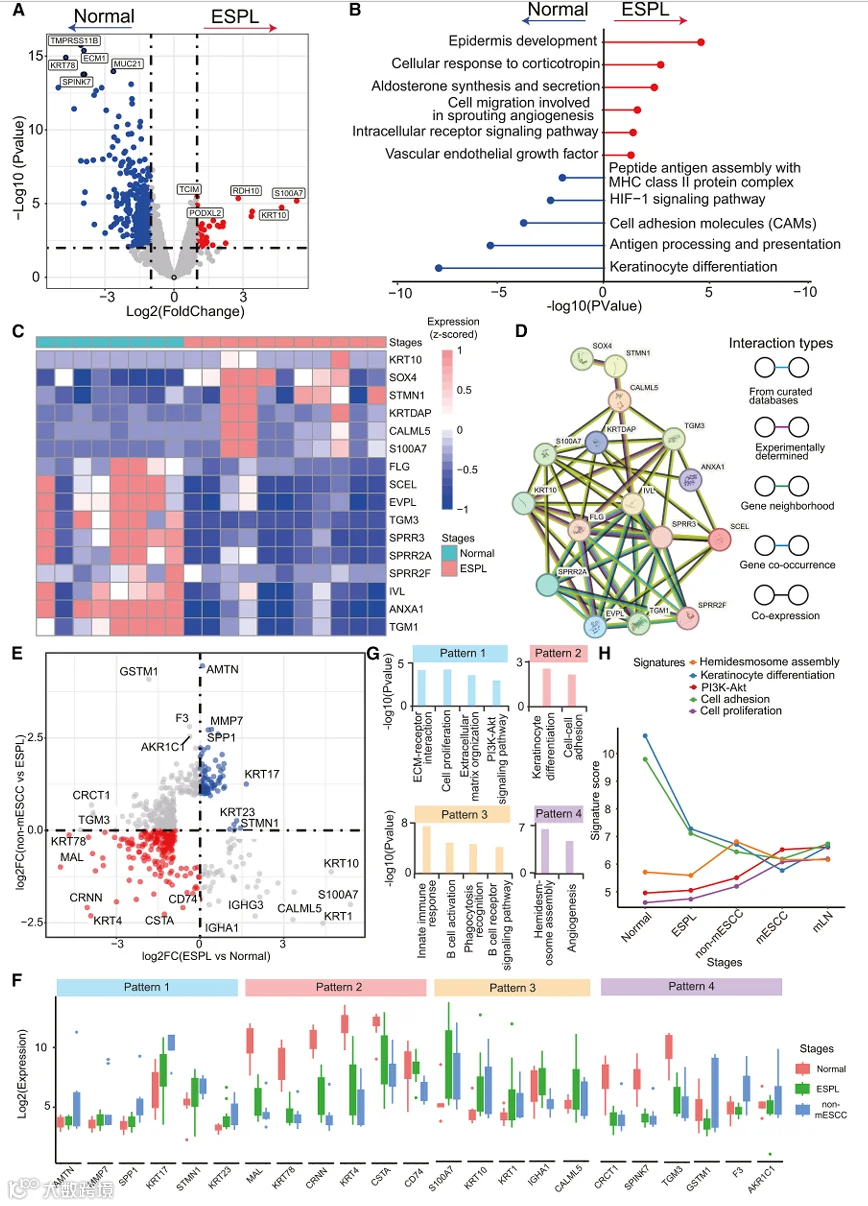
为了深入了解促进肿瘤逐步起始的机制,我们从转录模式、调控功能、细胞基因特征以及细胞组成等多个方面,表征了跨ESPL和ESCC进展阶段的肿瘤生态系统中的多样化细胞组分。我们首先研究了从NE到ESPL过程中EP细胞中改变的基因,发现包括角化细胞分化和表皮发育在内的几个ESCC特异性肿瘤发生通路失调(图2A和2B;表S3)。具体来说,与NE中的表达相比,角化细胞分化和缺氧诱导因子-1α信号通路等表达程序在ESPL中显示出显著降低(图2B)。此外,与角化细胞分化相关的基因如小富脯氨酸蛋白3(SPRR3)、胱抑素A(CSTA)和转谷氨酰胺酶1(TGM1)的表达在从NE到ESPL的进展中大幅下降,这与先前研究显示异常角化细胞分化是皮肤和食管鳞状细胞癌起始的关键因素的结果一致。相比之下,在ESPL中观察到表皮发育程序的表达显著增加,同时几个胚胎样基因如SRY相关HMG-box(SOX4)和角化细胞分化相关蛋白(KRTDAP)上调(图2C)。这些表达模式表明,表皮发育和角化细胞分化的协同调控在维持食管鳞状上皮的正常状态和促进癌前病变发生中起关键作用。结合先前发现以及表皮发育和角化细胞分化相关蛋白质之间相互作用的证据,我们推断角化细胞的去分化可能是ESPL发展中的关键过程,该过程由多个调控程序协调。此外,胚胎样基因的激活可能是角化细胞去分化的起始步骤,它进一步诱导表皮发育相关基因(如KRTDAP、钙调蛋白样蛋白5 [CALML5]、角蛋白10 [KRT10]和银屑病素 [S100A7]),导致角化细胞分化相关基因(如丝聚蛋白 [FLG]、小富脯氨酸蛋白2F [SPRR2F]、膜联蛋白A1 [ANXA1]和转谷氨酰胺酶3 [TGM3])的下调(图2D)。值得注意的是,最近的实验研究表明,Sox9的增加和ANXA1表达的降低对于食管癌起始至关重要,进一步证实了我们的发现。
我们接下来探索了可能驱动non-mESCC从ESPL演化的基因程序。通过同时分析三个阶段的失调基因,我们将差异基因分为四种表达模式(图2E和2F;表S3)。模式1中有36个基因,这些基因在NE中表达较低,在ESPL和non-mESCC中表达逐渐升高。如基因集富集分析所示(图2G),这些逐渐失调的基因显著富集于细胞外基质(ECM)-受体相互作用、PI3K/AKT信号通路和ECM组织过程,提示角蛋白17(KRT17)和分泌型磷蛋白1(SPP1)等基因的持续激活可能促进ESCC的发展。此外,基于模式2基因的功能富集,角化细胞分化过程在ESCC起始期间持续下降。在模式3中,参与先天免疫应答和B细胞活化正调控的基因在ESPL中高表达,但在non-mESCC中表达较低。这表明ESPL中的食管上皮细胞可能通过表达特定基因如免疫球蛋白重链恒定区α1(IGHA1)和IGHG3来激活先天免疫和B细胞介导的免疫应答,但这些基因随后在non-mESCC阶段被下调。最后,我们研究了模式4中在ESPL中下调但在non-mESCC中激活的基因。发现这些基因参与血管生成和半桥粒组装的调控。因此,我们得出结论,角化细胞分化和表皮发育的失调对于肿瘤起始至关重要,由胚胎样基因触发的角化细胞去分化在ESPL发展中起关键作用。此外,ECM-受体相互作用和PI3K/AKT信号通路的持续激活,以及动态的免疫应答变化,驱动了从ESPL到non-mESCC的进展。
为了进一步剖析空间上相邻的上皮和肿瘤微环境AOs是否存在功能关联,我们进行了靶向空间共定位分析。具体来说,我们筛选了EP区室与其相邻免疫区室在同一空间区域内共定位的AOs,重点关注ESCC的起始阶段。随后对上皮AOs的ESCC相关特征基因与相邻免疫区室中免疫调节相关基因进行相关性分析,揭示了上皮基因表达模式与免疫抑制特征之间的阶段特异性关联(图S2A-S2C)。值得注意的是,作为ECM-受体相互作用和PI3K/AKT信号关键介质的SPP1,在相邻MC区室中与免疫抑制性巨噬细胞标志物CD274显示出显著的正相关。与ESPL阶段相比,这种相关性在non-mESCC中进一步增强(图S2D),表明在ESCC起始过程中,上皮细胞中SPP1的表达与PD-L1阳性巨噬细胞之间存在潜在的空间协调的免疫抑制串扰。
接下来,我们对上皮区室中识别的不同基因表达模式所对应的通路富集结果在ESCC起始和进展的不同阶段进行了动态变化的系统分析。为了定量评估通路动态的活性,我们为每个通路定义了一个“特征评分”,该评分计算为该特定通路中富集的所有基因表达的中位数。我们的结果揭示了关键通路在ESCC发展过程中的不同轨迹模式(图2H)。与肿瘤发生和细胞增殖相关的通路,如PI3K/AKT信号通路和细胞增殖相关通路,其特征评分从正常组织阶段到晚期ESCC呈现出持续上升的趋势。这种进行性激活与模式1基因(例如,SPP1和KRT17)的逐渐上调一致,这些关键分子驱动了这些促肿瘤通路的激活。相反,与上皮稳态相关的通路,包括角化细胞分化通路(模式2的核心通路)和细胞粘附通路,在相同的疾病进展轨迹中显示出特征评分的稳步下降。这种持续的抑制反映了在恶性转化过程中正常上皮细胞身份的丧失。半桥粒组装通路显示出独特的双相动态:其特征评分从正常上皮到ESPL下降,但在晚期ESCC中反弹。总的来说,这些发现证明了在ESCC发展过程中,上皮分化程序的抑制与促肿瘤发生信号的激活之间存在功能权衡,进一步证实了驱动恶性进展的协调的分子重编程。
3.空间全转录组分析解析ESCC起始过程中间质微环境的变化
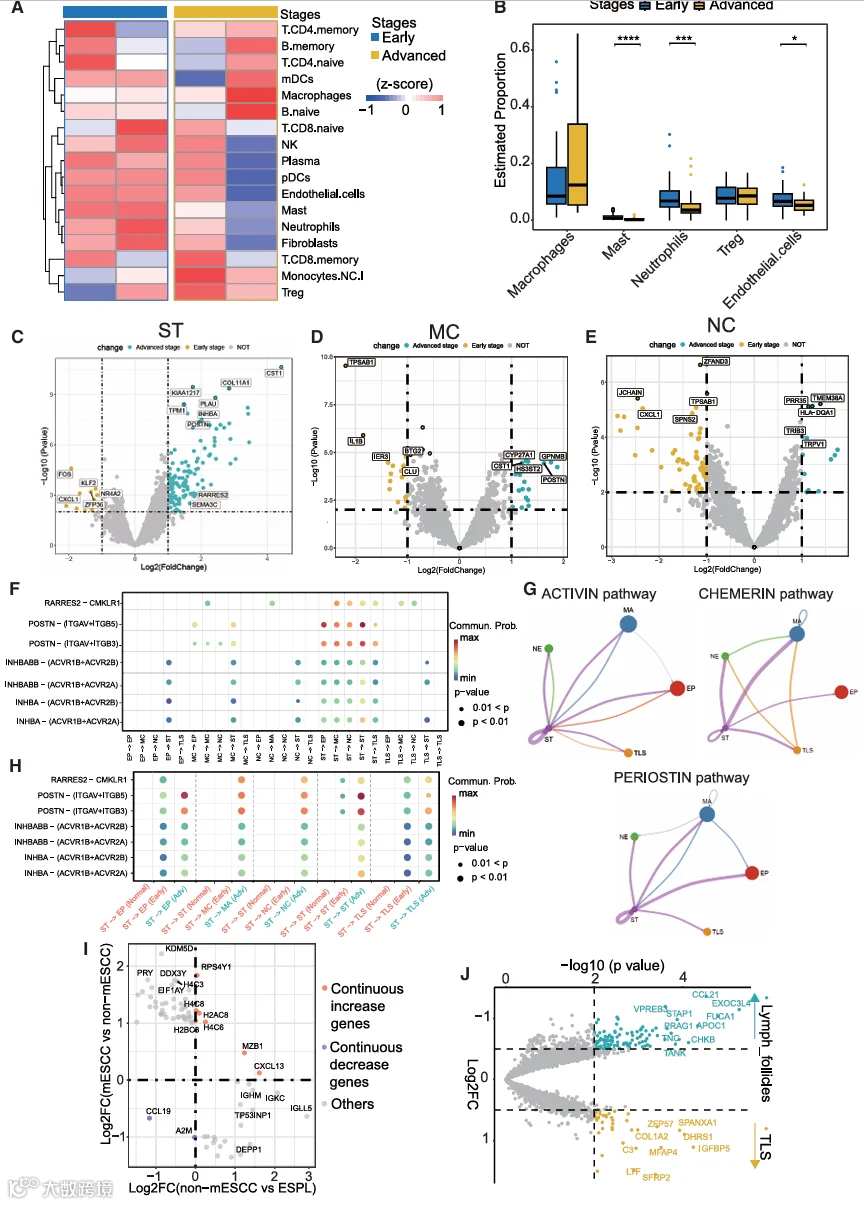
由于肿瘤起始和发展受到TME的强烈影响,我们进一步研究了免疫浸润以及免疫细胞的组成和状态,以探索ESCC起始过程中TME的变化。我们首先基于PanCK阴性区室的空间WTA数据识别并量化了免疫细胞。统计分析显示,与NE组织相比,ESPL中B细胞和浆细胞的比例显著增加(图3A、3B和3A;表S4),独立分析进一步验证了这一点(图3C)。值得注意的是,我们在ESPL中发现初始B细胞和记忆B细胞的水平均有所增加(图3C和S3B;表S4)。这些观察结果与上述关于我们研究中EP区室的发现以及先前的研究一致,即在ESPL阶段,先天免疫和B细胞介导的免疫应答被激活。此外,与ESPL阶段相比,non-mESCC中CD4+和CD8+ T细胞的比例显示出显著下降,表明淋巴细胞浸润减少可能促进ESCC起始(图3A-3C)。为了进一步证实这些发现,我们对从正常到non-mESCC阶段的这些样本进行了多重免疫荧光(mIF)染色(图S4A);从NE到ESPL阶段,CD20阳性细胞的百分比显著增加,而从ESPL到non-mESCC阶段,CD8+和CD4+ T细胞的百分比显著下降(图3D和3E)。
我们在本研究中采用的空间分割策略使我们能够进一步分析MC、NC和TLS区室中的分子变化。尽管在ESCC起始过程中未观察到MC比例的显著变化,但对其基因表达模式的深入分析揭示了多个与肿瘤发生相关的通路(图3F)。例如,在从ESPL到non-mESCC的进展过程中,MC中观察到补体激活和免疫应答的下调,以及细胞生长和增殖的上调,其中有多个基因参与这些过程的调控(图3F和3G)。此外,NC在ESPL中呈下降趋势,但在ESCC中增加(图3C)。参与调节细胞因子和凋亡过程的基因主要在NC中失调,表明它们在ESCC肿瘤发生中的调控作用(图3F和3G)。我们接下来使用解卷积方法对巨噬细胞和中性粒细胞进行了亚型特异性分析,以研究这些细胞亚型在ESCC起始和进展过程中的变化。我们的结果显示,促肿瘤的肿瘤相关中性粒细胞2(TN2)的比例从ESPL阶段到non-mESCC阶段显著增加,而肿瘤相关巨噬细胞2(TAM2)的比例在mLN阶段显著升高。这些亚型特异性变化进一步证实了巨噬细胞和中性粒细胞在ESCC起始和转移中发挥着阶段依赖性的功能作用(图3H)。最后,作为TME的重要组成部分,TLSs是主要由B细胞组成的异位淋巴样聚集体,在肿瘤适应性免疫中起关键作用。通过分析TLSs表达模式和调控通路的变化,我们发现与B细胞和免疫应答通路相关的几个基因在non-mESCC阶段的TLSs中失调,并且载脂蛋白B mRNA编辑酶催化亚基3A(APOBEC3A)基因表达显示出显著差异(图3F和3I)。为了进一步验证这些发现,使用独立样本进行了mIF。比较ESPL和non-mESCC中TLS区室的蛋白质信号显示,随着阶段进展,APOBEC3A的表达显著增加(图3J和3K),提示TLS中APOBEC3A的表达可能反映ESCC的起始。我们进一步分析了TLS内APOBEC3A表达与各种免疫细胞群的相关性,以及其与关键免疫调节基因的相关性。结果显示,APOBEC3A与浆细胞呈显著正相关(R = 0.77,p = 0.021);在免疫调节基因中,APOBEC3A也与IGHM(B细胞活化和抗体分泌的关键标志物)显示出最强的正相关(图S4B和S4C)。这些相关性发现为APOBEC3A的潜在调控机制提供了初步线索,提示它可能参与调节TLS内的B细胞/浆细胞功能。综上所述,这些结果表明,巨噬细胞、中性粒细胞和TLSs各自独特的调控功能为理解ESCC起始病理过程中不同免疫细胞类型的复杂交叉调控提供了基础性见解。
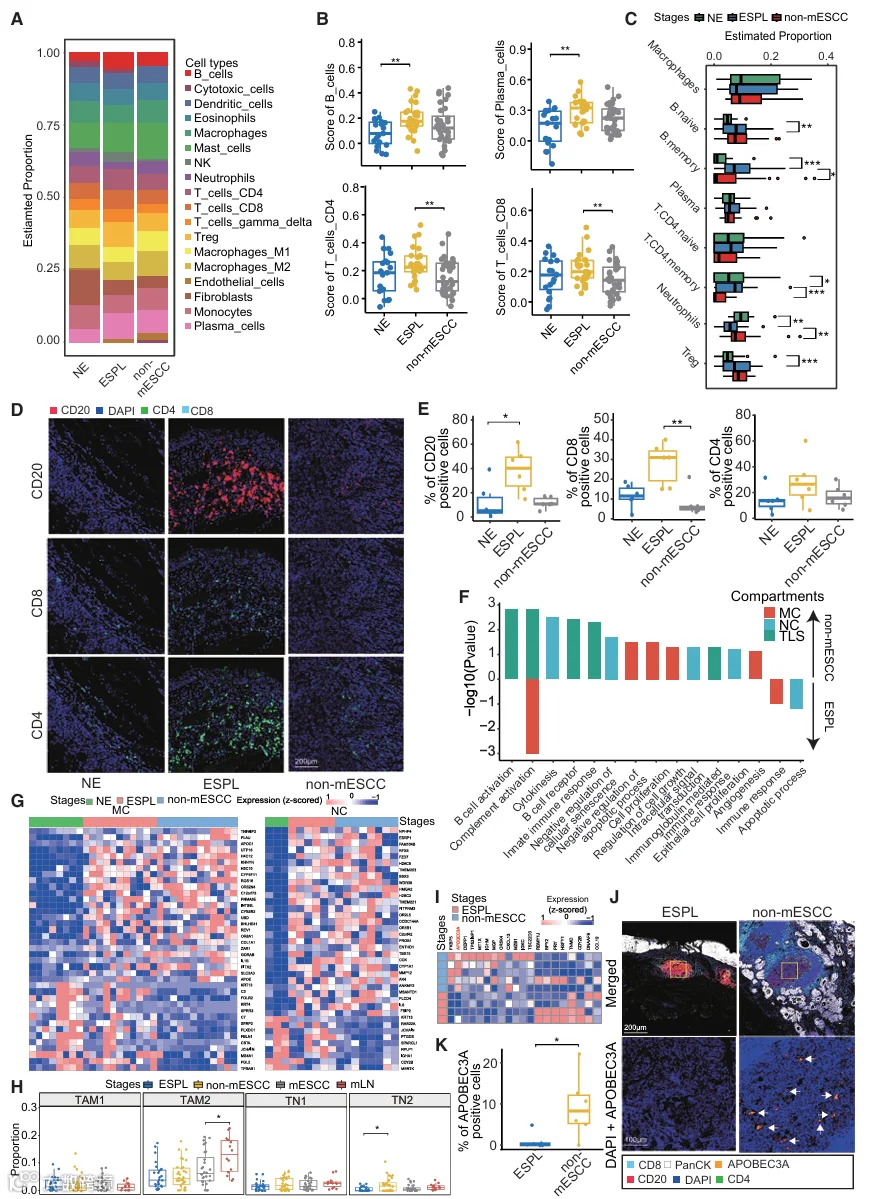

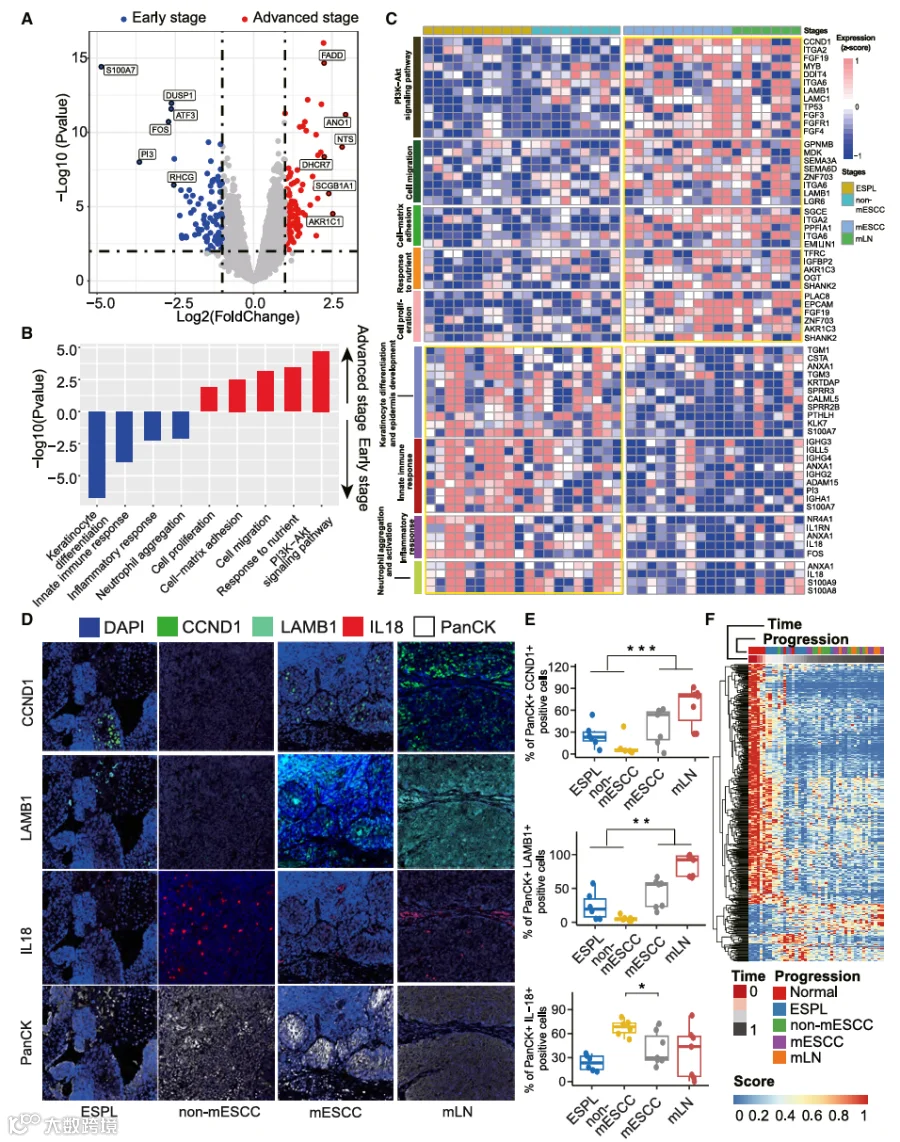
4.TME中的转录组学景观揭示从早期ESCC向晚期阶段过渡的特征
理解食管癌前病变如何演变为侵袭性或转移性肿瘤,对于改善ESCC预后可能具有深远意义。在本研究中,我们还旨在识别从早期阶段(ESPL和non-mESCC)到晚期阶段(mESCC和mLNs)ESCC过程中EP区室潜在的分子调控机制(图4A;表S3);几个与癌症进展相关的通路,如PI3K/AKT信号通路以及细胞迁移、粘附和增殖过程,在晚期肿瘤中显著上调(图4B)。值得注意的是,PI3K/AKT信号通路在肿瘤起始过程中持续上调,并且该通路中的其他基因,如细胞周期蛋白D1(CCND1)和层粘连蛋白亚基β-1(LAMB1),在晚期阶段进一步上调(图4C),表明该通路的持续激活对于ESCC的起始和发展至关重要。值得注意的是,与在肿瘤起始阶段激活的基因相比,某些与营养应答相关的基因,如OGT和TFRC,仅在晚期阶段被激活。此外,晚期阶段炎症和先天免疫应答的降低表明它们主要在肿瘤起始阶段被激活,但在肿瘤进展过程中被抑制。值得注意的是,参与中性粒细胞聚集和激活的基因,如白细胞介素(IL-18)基因,在晚期阶段被下调。基于mIF实验验证,我们确定来自PI3K/AKT通路的CCND1和LAMB1的蛋白质表达在肿瘤进展阶段显著增加(所有p < 0.01),而IL-18显著降低(p < 0.05)(图4D和4E)。
为了系统阐明肿瘤发展,我们随后进行了拟时间分析以模拟ESCC的演化轨迹。基于EP细胞的分析显示,它们的转变与ESCC的发展阶段高度相关(图4F)。然后,我们取晚期阶段特异性基因集和拟时间相关基因集的交集,得到九个重叠基因(图S5A)。我们进一步基于癌症基因组图谱(TCGA)-ESCC数据集评估了这些基因的表达谱与临床结局之间的关联,发现只有OGT与不良预后显著相关(图S5B)。此外,我们将OGT与其他已确定的阶段特异性标志物(例如SPP1和MMP9)进行比较以阐明其预后效用。结果显示,高OGT表达与患者较差的预后显著相关(风险比[HR] = 1.66,95%置信区间[CI] = 1.04-2.7,p = 0.034)(图S5C),表明高OGT表达可能是ESCC进展的独立风险因素。基于公共ESCC scRNA-seq数据集,我们确认OGT在ESCC晚期阶段的肿瘤细胞群中被特异性激活(图S5D)。为了探索OGT在ESCC之外的更广泛预后价值,我们根据OGT表达水平对其他TCGA肿瘤队列的患者进行了分层,并分析了他们的无病生存期(DFS)。研究结果表明,在几个TCGA队列中——包括子宫体子宫内膜癌(UCEC)、葡萄膜黑色素瘤(UVM)、肾乳头状细胞癌(KIRP)和间皮瘤(MESO)——高OGT表达的患者表现出显著缩短的DFS(图S5E)。这证实了OGT可以作为多种癌症类型DFS的预后标志物,突显了其作为通用预后因子的潜力。总的来说,这些发现确立了OGT作为晚期ESCC中具有深远功能和临床意义的因子,强调了进一步深入研究其驱动ESCC进展的生物学机制的必要性。
5.OGT在体内外均可增强ESCC细胞的迁移和侵袭能力
为了进一步阐明OGT在ESCC中的生物学功能,我们首先基于76对ESCC和邻近非癌组织进行了免疫组织化学分析(IHC)以检测OGT的表达水平(图5A)。结果显示,与邻近非癌组织相比,ESCC组织中OGT表达显著增加(图5B)。随后,对样本的临床和病理数据进行分析。观察到OGT表达水平与ESCC患者的病理T分期以及TNM分期之间存在统计学显著相关性,而OGT表达与患者的年龄、性别、吸烟史、饮酒史或肿瘤大小之间未发现显著相关性(表S5),表明OGT可能是参与ESCC进展的重要风险因素。
然后,我们通过RT-qPCR检测了正常食管HEEC细胞系和四种食管鳞状癌细胞系(KYSE30、KYSE150、KYSE410和KYSE450)中OGT基因的mRNA水平(图S6A)。接下来,建立了瞬时和稳定的过表达或敲低模型,同时过表达组进一步接受小分子OGT抑制剂OSMI-1(30 μM)处理,以初步评估OGT在ESCC细胞行为中的功能作用。通过蛋白质印迹(WB)分析确认了转染效率,同时也显示O-糖基化水平与OGT正相关,并且在OSMI-1处理后显著下调(图5C和5D)。通过平板克隆形成、细胞计数试剂盒-8(CCK8)、Transwell和伤口愈合实验评估的ESCC细胞恶性表型,在OGT敲低后显著受到抑制。相反,OGT过表达显著增强了这些恶性表型;然而,这种效应在OSMI-1处理后大大减弱(图5E-5H,S6B和S6C)。这些结果也证明了OGT介导的细胞运动性增强依赖于其酶活性。为了研究OGT在体内促进肿瘤生长和转移的作用,使用KYSE150和KYSE450细胞建立了两种类型的异种移植小鼠模型。在KYSE450皮下模型中,OGT敲低显著抑制了肿瘤生长,与对照组相比肿瘤体积显著缩小(图5I)。在肺转移模型中,注射后第7周,与对照组相比,oeOGT组小鼠表现出显著更强的肺部生物发光信号(图5J)、更多的表面结节(图5K)以及H&E切片上更大的转移面积(图5L),表明转移潜能增强。综上所述,来自体外和体内实验的证据表明,OGT在ESCC的进展和转移中发挥促进作用,凸显了其作为ESCC研究中关键分子靶点的潜力。
为了进一步研究OGT在ESCC中的分子机制,我们首先对OGT敲低和相应的对照细胞系进行了全转录组测序,鉴定出568个差异表达基因(DEGs),其中在OGT敲低组中375个上调,193个下调(图S6D;表S3)。同时,我们基于全转录组DSP数据构建了一个共表达网络,以筛选与OGT高度相关的基因,得到458个显著OGT相关的DEGs(图S6E)。富集分析显示,与角化细胞增殖、细胞粘附和凋亡过程相关的通路在OGT低表达组中被激活,而包括细胞周期、DNA复制以及肿瘤进展相关通路如Janus激酶-信号转导及转录激活因子(JAK-STAT)和Wnt信号通路在内的通路在OGT高表达组中显著富集,这些发现通过蛋白质组学分析得到了进一步验证(图5M)。考虑到OGT广泛参与蛋白质修饰,我们使用OGT过表达和Vector对照的ESCC细胞进行了定量O-GlcNAc糖蛋白组分析,这是一种基于质谱的方法,能够鉴定受OGT表达调节的O-GlcNAcylated底物。通过整合蛋白质组学和糖基化质谱结果,我们确认了OGT高表达组中细胞增殖相关通路的激活以及OGT低表达组中细胞凋亡通路的富集。基于流式细胞术的功能实验,我们观察到OGT敲低增加了KYSE450细胞的凋亡率,并改变了细胞周期分布,表现为G2/M期比例升高和S期比例降低(图5N,5O,S6F和S6G)。鉴于几个OGT相关通路也可能由组成型PI3K/AKT激活驱动,我们进一步研究了OGT与PI3K/AKT通路之间的功能性串扰关系。我们发现,OGT过表达诱导的O-GlcNAcylation增加与AKT Ser473磷酸化升高相关,而PI3K抑制剂LY294002或OGT抑制剂OSMI-1可有效抑制这种磷酸化。值得注意的是,在所有条件下,总PI3K和磷酸化PI3K、总AKT蛋白水平均未改变(图S6H)。这些结果表明,OGT通过PI3K下游的O-GlcNAcylation影响AKT磷酸化,可能影响包括JAK-STAT和Wnt信号在内的多个AKT依赖性通路。总之,这些发现证明OGT表达通过促进多条促肿瘤通路和抑制凋亡通路在ESCC进展中发挥关键作用,突出了OGT作为食管鳞状细胞癌潜在治疗靶点的价值。
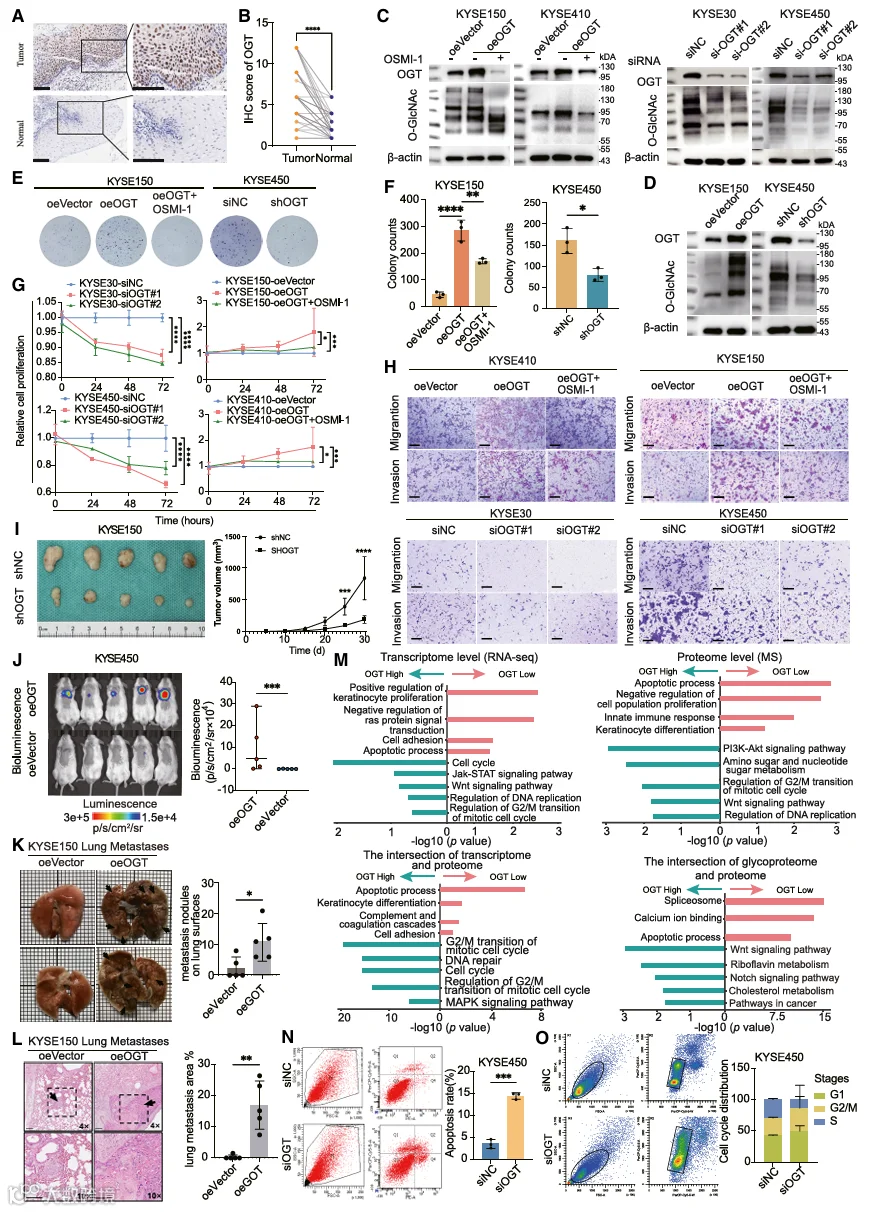
 我们接下来研究了ESCC进展过程中肿瘤生态系统重塑是如何发生的。不同细胞亚型比例的比较揭示了ESCC早期和晚期阶段细胞丰度的高度动态变化(图6A)。在晚期阶段观察到肥大细胞、中性粒细胞和内皮细胞大幅减少,而巨噬细胞呈增加趋势(图6B和S7A)。为了进一步理解ESCC进展过程中TME内的变化和相互作用,我们对ST、MC和NC区室进行了差异分析(图6C-6E;表S3)。基于各自DEGs的通路富集分析显示,在ST区室中,PI3K/AKT、血管生成和细胞迁移通路在晚期阶段更为活跃(图S7B)。在MC区室中,涉及中性粒细胞和T细胞的正向调控转向了晚期阶段增强的胆固醇代谢和细胞外空间活动(图S7B)。在NC区室中,尽管在早期阶段观察到免疫应答(如B细胞活化和白细胞迁移)的显著参与,但在ESCC的后期阶段观察到免疫活性受抑并转向表皮细胞分化和葡萄糖跨膜转运调控(图S7C)。此外,为了理解参与这些通路变化的DEGs如何在不同的区室中相互作用,我们在CellChat数据库中筛选了上述DEGs,识别出直接参与ESCC晚期阶段的细胞-细胞相互作用(CCI)通路(图6F;表S6)。这些不同区室之间的细胞-细胞相互作用通过ACTIVIN [INHBA->(ACVR1B+ACVR2)]、CHEMERIN (RARRES2->CMKLR1)和PERIOSTIN [POSTN->(ITGAV+ITGB5)] 通路发生(图6G)。对正常食管组织、早期ESCC和晚期ESCC样本中CCI通路的进一步分析揭示了关键信号轴在阶段特异性激活模式(图6H)。ACTIVIN、CHEMERIN和PERIOSTIN介导的相互作用在正常阶段未显示出显著的信号强度,而所有三条通路在早期ESCC中被激活,表现为ST与EP/TLS之间的细胞间串扰;其中,POSTN和ACTIVIN通路在晚期阶段显示出进一步增强的信号强度。值得注意的是,ST与MC/NC之间通过ACTIVIN、CHEMERIN和PERIOSTIN依赖的CCI仅在晚期ESCC中以显著增加的信号强度被检测到,并且在正常和早期阶段保持非活跃状态。此外,对免疫调节性CCI通路的评估显示,在整个ESCC进展过程中,TLS中CXCL13表达持续上调,并且CXCL13通过受体CXCR5、CXCR3和ACKR1与MC、NC和ST进行了强烈的细胞间通讯(图S7C)。这些发现共同证明了ESCC进展过程中CCI网络的进行性重塑,伴有阶段特异性诱导和增强的间质-上皮/TLS及间质-免疫相互作用。
最近的一项研究报告称,TLS变化在ESCC的发展中起重要作用。为了探索作为ESCC进展过程中TME关键部分的TLS的变化,我们比较了整个肿瘤进展过程中TLS中的DEGs。我们确定了三种不同的基因表达模式(图6I)。例如,趋化因子(C-X-C基序)配体13(CXCL13)、边缘区B和B1细胞特异性蛋白(MZB1)和H4簇状组蛋白6(H4C6)在肿瘤起始和进展过程中持续且显著过表达。CXCL13是参与招募B细胞和滤泡辅助性T(Tfh)细胞的主要趋化因子,验证了肿瘤进展过程中TLS的形成。相反,趋化因子(C-C基序)配体19(CCL19)和α-2-巨球蛋白(A2M)在此过程中被显著抑制。由于这些免疫相关基因在抑制肿瘤细胞迁移和转移中起关键作用,它们受抑的表达表明免疫细胞消除和抑制肿瘤转移的能力受损,导致晚期阶段迁移的可能性更高。TLS功能障碍相关基因与关键耗竭T细胞(Tex)标志物(例如,PDCD1、LAG3和TOX)之间的相关性分析揭示,在功能失调的TLS中表达上调的基因与Tex标志物PDCD1和TOX呈正相关(图S7D)。为了进一步剖析TLS功能障碍的潜在驱动因素,我们计算了每个TLS的功能障碍特征评分,以及它与晚期阶段EP和ST区室中失调信号通路特征评分的相关性。对同时包含EP和TLSs,或ST和TLS区室的样本进行分析显示,来自EP区室的失调通路与TLS功能障碍表现出相对较强的正相关,而来自ST区室的失调通路则显示出较弱或负相关(图S8A和S8B)。当聚焦于TLS区室时,我们观察到EP相关的“营养应答”特征评分与TLS功能障碍之间存在显著的正相关(R = 0.68,p = 0.014)(图S8A)。值得注意的是,与“营养应答”通路中的其他基因相比,TFRC和OGT基因显示出与TLS功能障碍最强的相关性(图S8C)。这些发现提示,在晚期ESCC期间,TLS功能障碍可能由肿瘤上皮区室中异常的“营养应答”信号通路驱动。
关于ESCC转移的过程,我们对TLSs和淋巴结内淋巴滤泡的DEGs分析揭示了值得注意的发现(图6J;表S3)。CCL21(一种招募T细胞和树突状细胞的趋化因子)和TRAF家族成员相关核因子κB(NF-κB)激活剂(TANK,一个参与NF-κB通路的基因)的显著增加,表明淋巴滤泡中免疫应答增强。然而,与肿瘤转移相关的基因如外泌体复合物组分3样蛋白4(EXOC3L4)和生腱蛋白-C(TNC)的过表达,表明mLN中的肿瘤细胞强烈抵抗免疫系统。相反,在TLSs中,虽然一些调节免疫功能的基因仍然显著升高,但与免疫细胞募集相关的细胞因子的表达受到抑制。这意味着在肿瘤进展的晚期阶段,TLSs无法招募新的免疫细胞,从而失去了有效抑制肿瘤细胞的能力。
我们接下来研究了ESCC进展过程中肿瘤生态系统重塑是如何发生的。不同细胞亚型比例的比较揭示了ESCC早期和晚期阶段细胞丰度的高度动态变化(图6A)。在晚期阶段观察到肥大细胞、中性粒细胞和内皮细胞大幅减少,而巨噬细胞呈增加趋势(图6B和S7A)。为了进一步理解ESCC进展过程中TME内的变化和相互作用,我们对ST、MC和NC区室进行了差异分析(图6C-6E;表S3)。基于各自DEGs的通路富集分析显示,在ST区室中,PI3K/AKT、血管生成和细胞迁移通路在晚期阶段更为活跃(图S7B)。在MC区室中,涉及中性粒细胞和T细胞的正向调控转向了晚期阶段增强的胆固醇代谢和细胞外空间活动(图S7B)。在NC区室中,尽管在早期阶段观察到免疫应答(如B细胞活化和白细胞迁移)的显著参与,但在ESCC的后期阶段观察到免疫活性受抑并转向表皮细胞分化和葡萄糖跨膜转运调控(图S7C)。此外,为了理解参与这些通路变化的DEGs如何在不同的区室中相互作用,我们在CellChat数据库中筛选了上述DEGs,识别出直接参与ESCC晚期阶段的细胞-细胞相互作用(CCI)通路(图6F;表S6)。这些不同区室之间的细胞-细胞相互作用通过ACTIVIN [INHBA->(ACVR1B+ACVR2)]、CHEMERIN (RARRES2->CMKLR1)和PERIOSTIN [POSTN->(ITGAV+ITGB5)] 通路发生(图6G)。对正常食管组织、早期ESCC和晚期ESCC样本中CCI通路的进一步分析揭示了关键信号轴在阶段特异性激活模式(图6H)。ACTIVIN、CHEMERIN和PERIOSTIN介导的相互作用在正常阶段未显示出显著的信号强度,而所有三条通路在早期ESCC中被激活,表现为ST与EP/TLS之间的细胞间串扰;其中,POSTN和ACTIVIN通路在晚期阶段显示出进一步增强的信号强度。值得注意的是,ST与MC/NC之间通过ACTIVIN、CHEMERIN和PERIOSTIN依赖的CCI仅在晚期ESCC中以显著增加的信号强度被检测到,并且在正常和早期阶段保持非活跃状态。此外,对免疫调节性CCI通路的评估显示,在整个ESCC进展过程中,TLS中CXCL13表达持续上调,并且CXCL13通过受体CXCR5、CXCR3和ACKR1与MC、NC和ST进行了强烈的细胞间通讯(图S7C)。这些发现共同证明了ESCC进展过程中CCI网络的进行性重塑,伴有阶段特异性诱导和增强的间质-上皮/TLS及间质-免疫相互作用。
最近的一项研究报告称,TLS变化在ESCC的发展中起重要作用。为了探索作为ESCC进展过程中TME关键部分的TLS的变化,我们比较了整个肿瘤进展过程中TLS中的DEGs。我们确定了三种不同的基因表达模式(图6I)。例如,趋化因子(C-X-C基序)配体13(CXCL13)、边缘区B和B1细胞特异性蛋白(MZB1)和H4簇状组蛋白6(H4C6)在肿瘤起始和进展过程中持续且显著过表达。CXCL13是参与招募B细胞和滤泡辅助性T(Tfh)细胞的主要趋化因子,验证了肿瘤进展过程中TLS的形成。相反,趋化因子(C-C基序)配体19(CCL19)和α-2-巨球蛋白(A2M)在此过程中被显著抑制。由于这些免疫相关基因在抑制肿瘤细胞迁移和转移中起关键作用,它们受抑的表达表明免疫细胞消除和抑制肿瘤转移的能力受损,导致晚期阶段迁移的可能性更高。TLS功能障碍相关基因与关键耗竭T细胞(Tex)标志物(例如,PDCD1、LAG3和TOX)之间的相关性分析揭示,在功能失调的TLS中表达上调的基因与Tex标志物PDCD1和TOX呈正相关(图S7D)。为了进一步剖析TLS功能障碍的潜在驱动因素,我们计算了每个TLS的功能障碍特征评分,以及它与晚期阶段EP和ST区室中失调信号通路特征评分的相关性。对同时包含EP和TLSs,或ST和TLS区室的样本进行分析显示,来自EP区室的失调通路与TLS功能障碍表现出相对较强的正相关,而来自ST区室的失调通路则显示出较弱或负相关(图S8A和S8B)。当聚焦于TLS区室时,我们观察到EP相关的“营养应答”特征评分与TLS功能障碍之间存在显著的正相关(R = 0.68,p = 0.014)(图S8A)。值得注意的是,与“营养应答”通路中的其他基因相比,TFRC和OGT基因显示出与TLS功能障碍最强的相关性(图S8C)。这些发现提示,在晚期ESCC期间,TLS功能障碍可能由肿瘤上皮区室中异常的“营养应答”信号通路驱动。
关于ESCC转移的过程,我们对TLSs和淋巴结内淋巴滤泡的DEGs分析揭示了值得注意的发现(图6J;表S3)。CCL21(一种招募T细胞和树突状细胞的趋化因子)和TRAF家族成员相关核因子κB(NF-κB)激活剂(TANK,一个参与NF-κB通路的基因)的显著增加,表明淋巴滤泡中免疫应答增强。然而,与肿瘤转移相关的基因如外泌体复合物组分3样蛋白4(EXOC3L4)和生腱蛋白-C(TNC)的过表达,表明mLN中的肿瘤细胞强烈抵抗免疫系统。相反,在TLSs中,虽然一些调节免疫功能的基因仍然显著升高,但与免疫细胞募集相关的细胞因子的表达受到抑制。这意味着在肿瘤进展的晚期阶段,TLSs无法招募新的免疫细胞,从而失去了有效抑制肿瘤细胞的能力。
更多结果和补充图表:doi:10.1016/j.xcrm.2026.102650
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!