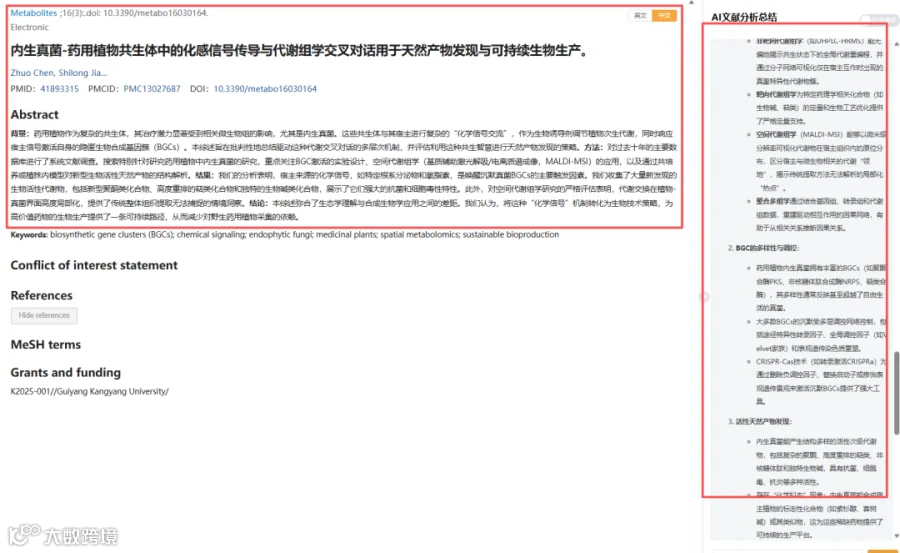
药用植物与内生真菌的共生关系构成了一个精妙的“化学信号”对话系统。本研究系统综述了利用多组学技术(代谢组学、空间代谢组学、基因组学、转录组学)解码这一共生机制的最新进展。研究发现,宿主植物的化学信号(如根系分泌物、oxylipins)可激活真菌中沉默的生物合成基因簇(BGCs),释放出大量新型聚酮、萜类及生物碱等活性分子。空间代谢组学更揭示,代谢交换高度集中于植物-真菌界面,而传统提取方法难以捕捉这种时空特异性。整合多组学与合成生物学策略,可将内生真菌开发为“微生物工厂”,实现高值药物的可持续生物制造,为替代野生采挖提供绿色解决方案。
今天给大家解读一篇2月发表在《Metabolites》上的题目为“Chemical Signaling and Metabolomic Crosstalk in Endophytic Fungi-Medicinal Plant Symbioses for Natural Product Discovery and Sustainable Bioproduction.”的文章。本文是一篇系统性综述,旨在批判性地总结驱动药用植物-内生真菌共生体内代谢交叉对话的多层机制,并评估利用这种共生“智能”进行天然产物发现的策略。文章通过综述近十年的研究,涵盖了从先进的代谢组学方法、真菌BGC调控网络、新发现的活性天然产物,到化学信号传导机制以及可持续生物生产的转化应用与挑战。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!


团队成员合影(位于上海陆家嘴中心,可随时预约参观)
题目:《内生真菌-药用植物共生体中的化感信号传导与代谢组学交叉对话用于天然产物发现与可持续生物生产》Chemical Signaling and Metabolomic Crosstalk in Endophytic Fungi-Medicinal Plant Symbioses for Natural Product Discovery and Sustainable Bioproduction
发表期刊:Metabolites
影响因子:3.7
研究背景:
药用植物是现代药物发现不可或缺的来源,但其可持续供应因过度采挖、生境丧失和天然代谢物产量低而受到威胁。内生真菌已成为药物相关化合物的一个有前途的替代来源。然而,基因组分析表明,内生真菌中大部分生物合成基因簇(BGCs)在标准实验室条件下处于沉默状态,这意味着其巨大的代谢潜力未被充分利用。解锁这种“隐匿”的多样性需要解析植物-内生真菌全息生物中亲密的分子对话,而多组学和生物技术的进步为此提供了可能。
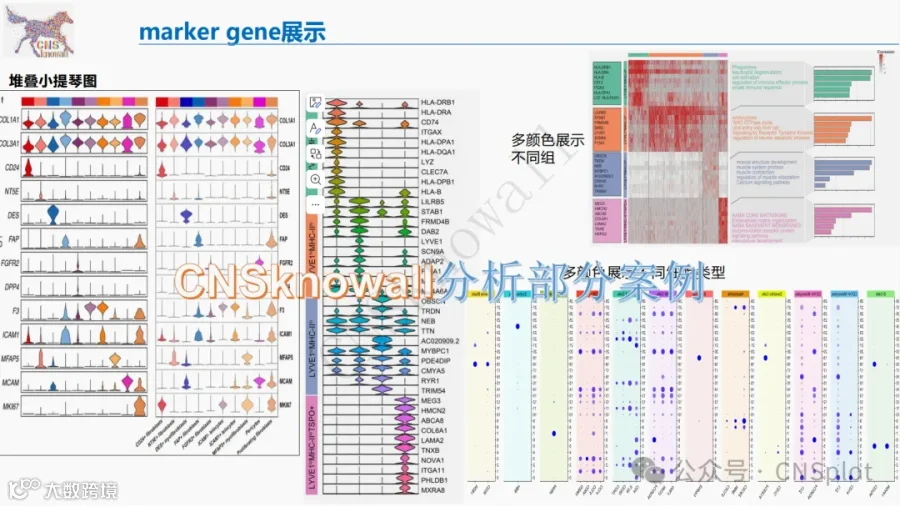
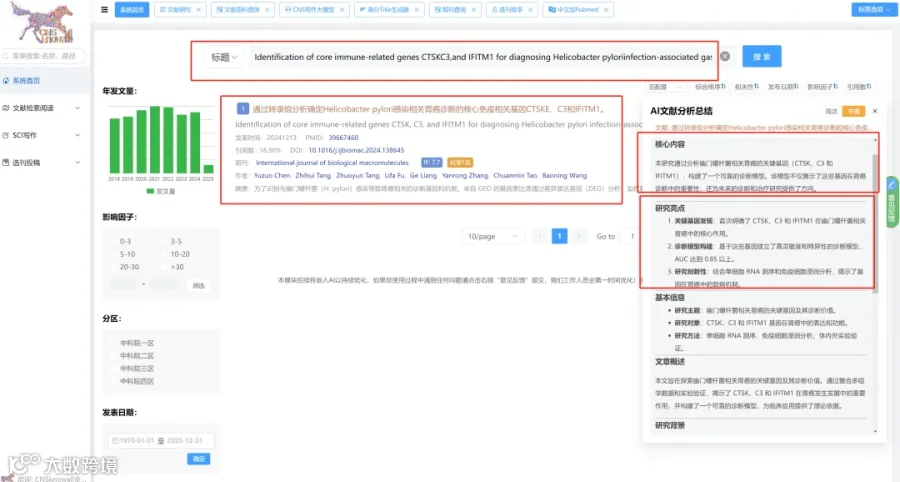
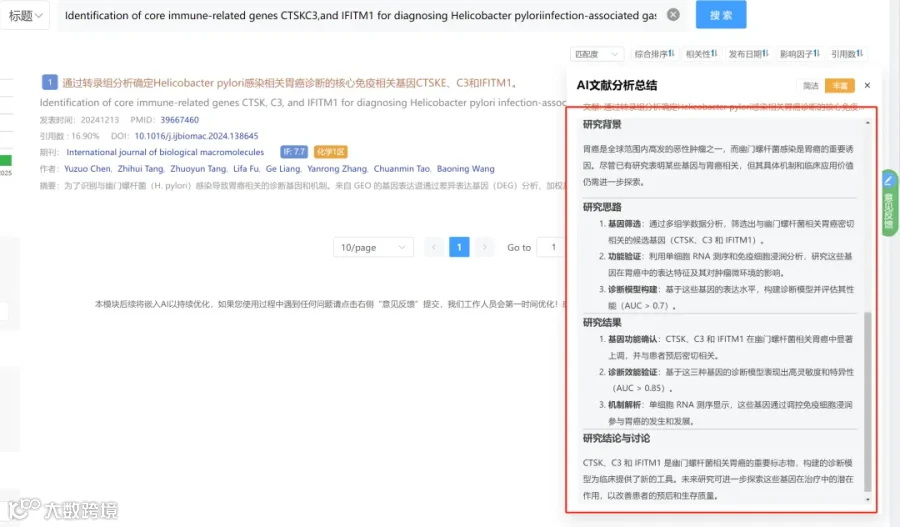
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
- 代谢组学策略
概述从无菌培养到空间原位模型等先进的代谢组学策略,用于破译共生相互作用。
- BGC与调控网络
详细阐述调控真菌BGCs的分子网络(如转录因子、表观遗传)及其激活策略(如CRISPR)。
- 活性天然产物
重点介绍通过共生相互作用发现的关键药理活性天然产物及其构效关系。
- 化学信号传导
合成化学信号传导的机制流,阐释信号分子如何介导定殖、调控宿主代谢并驱动共进化。
- 转化应用
讨论将上述机制转化为生物技术策略所面临的挑战,及其在可持续生物生产中的应用前景。
研究亮点:
- 揭示双向化学对话机制
明确了植物与内生真菌之间的化学信号传导是双向的。宿主化学信号可激发真菌沉默BGCs,而真菌产生的代谢物又能作为激发子,系统性地上调宿主植物的防御与药用成分合成通路。
- 强调空间代谢组学的关键作用
指出基质辅助激光解吸/电离质谱成像(MALDI-MSI)等空间代谢组学技术能可视化代谢物在植物-真菌界面的高度局部化分布,这是传统整体组织提取无法捕捉的关键生态信息。
- 提出从生态理解到合成应用的转化路径
文章不仅总结了机制,更构建了一个从生态洞察到生物技术应用的转化路线图,包括利用人工智能挖掘沉默BGCs、设计合成微生物群落、以及在工业底盘中进行异源表达,以替代对野生药用植物的采伐。
研究结果:
代谢组学策略:
- 非靶向代谢组学
(如UHPLC-HRMS)能无偏地揭示共生状态下的全局代谢重编程,并通过分子网络可视化仅在宿主互作时出现的真菌特异性代谢物簇。
- 靶向代谢组学
为特定药理学相关化合物(如生物碱、萜类)的定量和生物工艺优化提供了严格定量支持。
- 空间代谢组学
(MALDI-MSI)能够以微米级分辨率可视化代谢物在宿主组织内的原位分布,区分宿主与微生物相关的代谢“领地”,揭示传统提取方法无法解析的局部化“热点”。
- 整合多组学
通过结合基因组、转录组和代谢组数据,重建驱动相互作用的因果网络,有助于从相关关系推断因果关系。
BGC的多样性与调控:
-
药用植物内生真菌拥有丰富的BGCs(如聚酮合酶PKS、非核糖体肽合成酶NRPS、萜类合酶),其多样性通常反映甚至超越了自由生活的真菌。
-
大多数BGCs的沉默受多层调控网络控制,包括途径特异性转录因子、全局调控因子(如Velvet家族)和表观遗传染色质重塑。
-
CRISPR-Cas技术(如转录激活CRISPRa)为通过删除负调控因子、替换启动子或修饰表观遗传景观来激活沉默BGCs提供了强大工具。
活性天然产物发现:
-
内生真菌能产生结构多样的活性次级代谢物,包括复杂的聚酮、高度重排的萜类、非核糖体肽和独特生物碱,具有抗菌、细胞毒、抗炎等多种活性。
-
存在“化学拟态”现象:内生真菌能合成宿主植物的标志性化合物(如紫杉醇、喜树碱)或其类似物,这为这些稀缺药物提供了可持续的生产平台。
-
特定的支架变化、绝对构型和官能团修饰决定了代谢物的靶点选择性和药理安全性,为后续先导化合物优化提供了依据。
化学信号传导机制:
-
共生由复杂的信号网络(根系分泌物、植物激素、效应蛋白)驱动。植物激素(如水杨酸、茉莉酸)可作为转录触发器,直接激活真菌的沉默BGCs。
-
内生真菌定殖会重编程宿主初级代谢(如碳氮通量),为次级代谢物的生物合成提供前体,同时也能系统性上调宿主自身的防御和药用成分合成通路。
-
代谢物交换是高度局部化的,存在从紧密的代谢通道到选择性前体供给的相互作用谱系,共同维持全息生物的稳定性和化学复杂性。
-
宿主特异性主要由宿主驱动的代谢过滤(如根系分泌物)和共同进化塑造,基因组可塑性在定义宿主范围和生活方式中起核心作用。
研究总结:
- 结论
药用植物-内生真菌共生体是一个在化学层面深度整合的系统,其“化学信号”传导机制是连接生态理解与合成生物学应用的桥梁。将这种机制转化为生物技术策略,为高价值药物的生物生产提供了一条可持续途径,有助于减少对野生药用植物的依赖。
- 转化路线图
未来的发展需遵循从机会性分离转向精准发现和稳健部署的统一轨迹。这包括利用人工智能和多组学挖掘未充分采样的宿主谱系及极端环境中的BGCs;通过宿主-基因型-环境匹配重新设计接种剂管道;以及利用合成生物学工具构建不依赖宿主信号的工业化生产底盘。
- 挑战与前景
主要挑战包括难以培养的微生物、发酵过程中的产量衰减以及跨生产规模的代谢谱可重复性问题。人工智能(如AlphaFold2)在预测BGC酶结构、从而推断代谢物结构方面的应用,预示着将从随机筛选转向理性的、结构导向的发现新时代。最终,实现这些技术的可持续规模化需要标准化的功效协议和公平的菌种资源获取。
结果译文:
代谢组学通过捕捉初级和次生代谢的实时变化,为全生物体提供了明确的功能性读出的信息。与预测潜在功能的基因组学或转录组学不同,代谢组学直接将化学表型与内生菌定植、胁迫响应和治疗特性联系起来。基于非靶向质谱(MS)的工作流程,特别是与分子网络联用时,能够实现对无菌培养、共培养和活体植物条件下代谢变化的系统级映射。这些方法对于揭示沉默生物合成通路的激活和新代谢产物家族的出现至关重要。全局分析可通过靶向检测进一步补充,以量化特定的防御代谢物和共生生物标志物。将这些化学数据集与生态模型相结合,正越来越多地揭示特定化合物类别(如硫代葡萄糖苷和聚酮)如何驱动内生菌群落组装和宿主重编程。
非靶向代谢组学是探索药用植物-内生菌相互作用中尚未被充分表征的化学空间的主要策略。通过避免先验选择,高分辨液相色谱-串联质谱(LC-MS/MS)和核磁共振(NMR)平台能够对非接种与接种宿主,以及无菌培养与共培养系统进行无偏比较。这种比较方法将宿主来源分子与微生物产物区分开来,揭示了与促生长和药效相关化合物诱导相关的代谢重编程。
近期的应用证明了这一工作流程的强大功能。例如,对药用植物细胞悬浮液进行超高效液相色谱-高分辨质谱(UHPLC-HRMS)分析表明,内生菌接种刺激了先前未检测到的酚类和萘醌衍生物的积累。此外,分子网络已被证明对于可视化仅在宿主相互作用中出现的内生菌特异性代谢物簇至关重要,这表明了沉默基因簇的唤醒。在对辣根根部的研究中,非靶向LC-MS/MS结合扩增子测序揭示,特定代谢物类别的变化与真菌群落结构显著相关,突显了植物化学在过滤微生物组中的作用。类似地,基于NMR的Combretum lanceolatum分析鉴定了内生菌诱导的初级代谢变化,该变化将碳流转向防御相关前体。这些基础代谢组学实验设计的比较,包括其生物学背景和主要应用,总结于表1中。
虽然非靶向工作流程推动发现,但靶向代谢组学提供了验证药理相关化合物所必需的定量严谨性。通过采用优化的多反应监测转换和真实标准品,该方法提供高灵敏度和绝对定量,使研究人员能够严格地将内生菌定植方案或培养条件与特定生物碱、萜类和酚类物质的产量联系起来。这种定量精度对于机理研究和基于内生菌生产的生物工艺优化至关重要。“广泛靶向”代谢组学的发展现在允许对数百种已知代谢物进行高通量定量,为质量控制提供了可扩展的模板。例如,此类平台已成功绘制了药用植物(如橡胶草)中组织特异性积累图谱,为检测内生菌定植组织中的类似代谢变化提供了路线图。
在全生物体的特定背景下,靶向策略发挥着关键作用:确认内生菌来源的植物药物类似物、监测毒性标志物以及验证通过非靶向筛选优先发现的生物活性物质。一个显著的应用涉及青蒿内生菌,其中多变量分析优先筛选了醌类衍生物,随后通过靶向分离鉴定出大黄素和大黄酚甲醚,它们具有亚微摩尔IC₅₀值的强效抗疟活性。此外,靶向分析可以通过量化治疗后内源性代谢物的变化来解码草药的药效动力学。随着分析库的扩展,这些策略将对于标准化基于内生菌的植物药,以及将生态相互作用转化为可量化的药理学产出至关重要。
1.3. 空间代谢组学用于绘制宿主组织内代谢物分布图
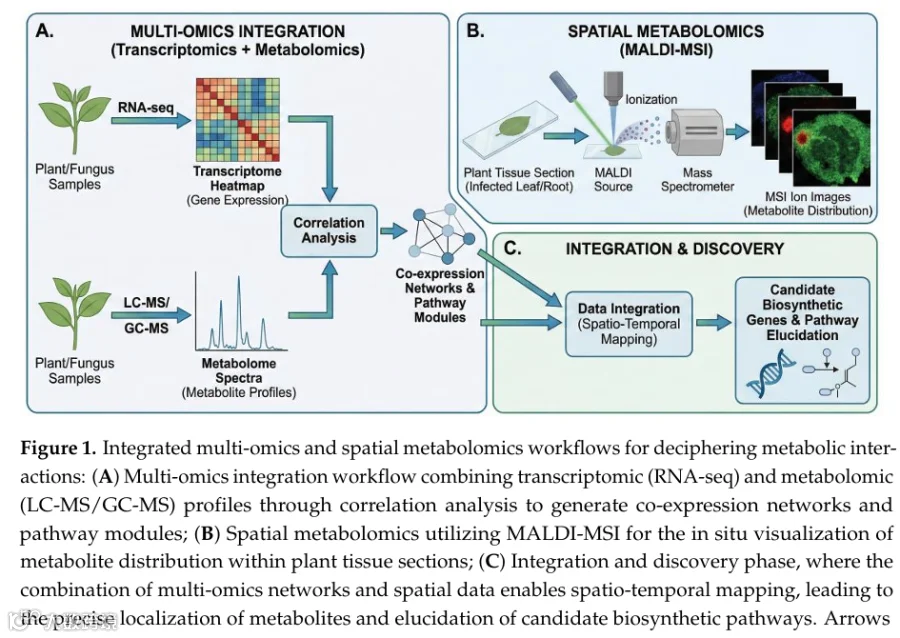
空间代谢组学,特别是基质辅助激光解吸电离质谱成像(MALDI-MSI),通过以微米级分辨率原位可视化代谢物分布,弥补了化学分析与组织学之间的差距。与整体提取不同,MSI重建离子图,以揭示脂质、次生代谢物和植保素在组织内的精确积累。最近的进展已将空间分辨率推至亚细胞水平(< 5 μm),并引入了多模态工作流程——例如将MSI与荧光原位杂交联用——以实现代谢物向特定微生物细胞或宿主结构的精确分配。值得注意的是,虽然MSI已在明确定义的植物-病原体模型(如可视化感染期间的真菌毒力因子)中得到广泛验证,但其在互惠性植物-内生菌共生中的应用是一个快速发展的前沿领域。在内生菌研究中,研究人员必须谨慎设计MSI工作流程,以区分防御性化学战与合作的共生化学交换。
在全生物体背景下,MSI对于区分宿主来源与微生物来源的代谢区域至关重要。对玉米、葡萄和水稻的研究已成功区分了病原体来源的毒素与局部的宿主防御“热点”,而整体分析无法解析这些热点。除了绘图之外,MSI还支持功能靶点验证;例如,在稻瘟病菌感染的大麦中可视化真菌黑色素通路的破坏,直接将代谢抑制与致病性降低联系起来。这些空间分辨的特征为整体代谢组学提供了机制性的补充层。
尽管具有这些能力,挑战依然存在。空间分辨率与灵敏度之间始终存在权衡,即使使用先进的离子化技术,也常常限制了低丰度次生代谢物的检测。此外,样品制备构成了主要瓶颈;植物组织的高含水量可能导致切片过程中代谢物的迁移,因此需要标准化方案以最小化脆弱的根-微生物界面的伪影。如图1所示,将这种高分辨率空间成像(MALDI-MSI)与转录组学-代谢组学相关分析整合,提供了一个全面的工作流程:它能够精确定位代谢物并原位鉴定其生物合成途径,从而解析全生物体的时空逻辑。
整合多组学结合了基因组学、转录组学和代谢组学数据,以重建驱动宿主-内生菌相互作用的因果网络。通过联合分析这些数据层,研究人员可以超越描述性基因列表,识别关键调控节点。例如,茶树和大麦中的共表达网络已成功将丝裂原活化蛋白激酶(MAPK)信号级联与防御相关的代谢模块联系起来,而真菌病原体中的代谢物-全基因组关联研究(GWAS)已将调控多态性与毒力因子关联起来。
解析这些复杂性状需要一个稳健的框架,其中用于BGC注释的高质量基因组组装与时间分辨的双RNA测序(dual RNA-seq)相结合,以捕获动态转录模块。然后将该转录组景观与匹配的代谢组学分析(特别是靶向中央碳通量和特异性防御化合物)相结合。随后的基于网络的整合——例如将加权基因共表达网络与代谢物丰度叠加——可以优先筛选候选酶用于功能验证。
然而,瓶颈依然存在。首先,全生物体巨大的化学多样性限制了代谢组的覆盖范围,常常导致系统性假阴性。其次,由于谱库不完整,只有一小部分检测到的特征能够被可靠地鉴定,存在明显的“注释鸿沟”。最后,跨不同空间尺度(单细胞MSI vs. 整体RNA-seq)的数据整合给稳健的网络分析带来了巨大的计算挑战。
1.5. 用于内生菌-宿主代谢组数据的生物信息学工具和工作流程
处理双生物数据集需要强大的生物信息学工作流程,能够解析复杂的宿主-微生物信号。完整的流程通常包括三个阶段:自动化预处理、统计功能分析和整合性通路重建。
预处理和分子网络。 对于非靶向LC-MS/MS数据,自动化流程(如UmetaFlow和MetMiner)现已提供从特征检测到对齐的端到端解决方案,并直接连接到全球天然产物社会分子网络(GNPS)。这种整合对于分子网络至关重要,分子网络通过聚类结构相关的代谢物,以直观地区分内生菌特异性类似物与宿主背景。例如,在Astrocaryum-内生菌系统中,此类工作流程成功分离了宿主富集和微生物富集的代谢物类别,从而能够在全生物体内追踪不同的化学谱系。对于基于NMR的分析,类似的多变量工作流程有助于分箱和谱库匹配,以跟踪定植过程中的初级代谢变化。
统计与功能解释。 下游分析越来越多地集中在基于网页的套件中,如MetaboAnalyst 4.0和XCMS Online。除了标准的单变量/多变量统计外,这些平台还提供联合通路分析模块,允许研究人员将基因组或蛋白质组数据直接叠加到代谢组失调图上。这些系统生物学能力对于将特定的代谢特征(如上调的防御化合物)与表型性状(如胁迫耐受性或抗病性)联系起来至关重要。
通路重建与整合。 为了从相关性走向因果关系,专门用于模拟代谢交接的工具正在涌现。MEANtools采用基于反应规则的算法,将质谱特征与转录组数据联系起来,从头重建候选生物合成途径。与此同时,像METABOLIC这样的基因组规模流程可从宏基因组数据中分析功能网络,以预测代谢物交换潜力。将这些推断的网络与测量的代谢组变化相结合,为破译植物-内生菌对话的“化学语言”提供了一个连贯的框架。
虽然这些计算工具使代谢组学普及化,但对于植物-内生菌研究来说,选择合适的流程仍然至关重要。分子网络对于聚类结构相关的类似物和通过谱库匹配识别已知化合物家族非常有效,但它常常难以处理内生真菌特有的全新骨架的“暗物质”。相比之下,基于碎片树的方法擅长从头预测未知代谢物的分子式和亚结构,不依赖于谱库。研究人员不应不加选择地应用生物信息学工具,而应遵循逻辑决策树:利用GNPS主要进行已知骨架的快速去重复,同时保留靶向MS/MS分子网络和像antiSMASH这样的计算机BGC预测工具,用于优先筛选新的或高度分化的BGC产物。因此,一个混合工作流程——结合使用GNPS进行宿主植物代谢物的快速去重复,以及使用SIRIUS/CANOPUS进行新型真菌天然产物的结构解析——代表了剖析这些共生系统中复杂化学相互作用的当前黄金标准。
代谢组学策略的机制性总结。 从基础的无菌筛选到整合多组学和MSI,提供了一个渐进的工作流程。非靶向代谢组学生成初始的整体图谱,多组学将这些表达图谱与其潜在的遗传能力(BGCs)联系起来,而空间代谢组学则原位可视化了其生态现实。
内生真菌拥有一个庞大的BGCs库,这些基因簇编码核心骨架形成酶——如非核糖体肽合成酶(NRPS)、聚酮合酶(PKSs)和萜烯合酶——以及必要的修饰酶和转运蛋白。大规模基因组挖掘已揭示了数十万个此类BGCs,其中绝大多数尚未被表征,突显了内生层中巨大的隐存化学多样性。然而,大多数基因簇在标准实验室条件下是转录沉默的。它们的激活受复杂调控网络的控制,涉及通路特异性转录因子、全局调控因子、染色质重塑和环境信号。
近期来自模式丝状真菌的见解说明了在内生菌中预期的调控结构。基因共表达网络已成功预测了控制多个BGCs的反式作用转录因子,包括那些缺乏明显内部调控因子的基因簇。这些因子的实验性过表达通常能调节多种次生代谢产物的产生,揭示了簇内与全基因组调控之间的广泛联系。此外,像Velvet家族的全局调控因子已被证明能协调影响分生孢子形成、胁迫响应和聚酮生物合成,从而广泛重塑次生代谢产物谱。这些层次与表观遗传机制相交,其中组蛋白修饰决定染色质可及性;对此机制的靶向扰动可以有效激活沉默的BGCs并使代谢产物输出多样化。进化分析进一步表明,BGC调控具有高度可塑性,转录因子常常发生分化或在相关物种中被重新用于控制不同的基因簇。
药用植物的内生菌拥有多样化的生物合成能力,这反映并常常延伸了自由生活真菌的能力。对单个分离株的基因组挖掘通常揭示每个基因组中数十个基因簇,以I型聚酮合酶(T1 PKS)、非核糖体肽合成酶(NRPS)、萜烯以及PKS-NRPS杂合系统为主。例如,来自莲雾的单个Talaromyces sp. 编码了76个预测的BGCs,包括罕见的膦酸酯和β-内酯簇,而来自岩白菜的Helotiales内生菌包含超过77个簇,涵盖PKS、NRPS和核糖体合成肽(RiPP)家族。
对这些BGCs进行分类揭示了由宿主关联塑造的独特酶学特征。PKS簇常利用还原型和非还原型T1 PKS生成蒽醌等骨架;在金丝桃属植物来源的内生菌中,特定的非还原型PKS簇已与金丝桃素样生物合成相关联。NRPS和杂合PKS-NRPS簇对肽类多样性有显著贡献,驱动了真菌谱系中二酮哌嗪和复杂杂合骨架的产生。萜烯和梅罗萜类簇也广泛存在,通常嵌入能产生植物相关挥发性物质(如芳樟醇)的萜烯合酶基因,或利用不寻常的异戊烯基转移酶-环化酶逻辑生成聚酮-萜烯杂合体。虽然大规模基因组学表明大多数BGCs属于分类学上受限的基因簇家族,但内生菌似乎将这些核心模块组装成独特的库,例如在金丝桃属植物共生体中发现的独特蒽醌相关簇,与岩白菜内生菌中梅罗萜类富集的谱形成对比。
2.2. 用于预测生物活性途径的基因组挖掘和计算机模拟预测
系统性的基因组挖掘彻底改变了在药用植物内生菌中发现潜在生物合成能力的方式。在基因组水平上,像antiSMASH这样的计算平台现在能够快速识别和基于规则地注释BGCs,提供与实验表征途径的簇级比较。
近期的应用展示了这一方法的强大功能。对来自莲雾的内生菌Talaromyces sp. 进行全基因组测序,揭示了76个BGCs——涵盖聚酮、非核糖体肽和萜烯家族——并允许将簇特异性分配到如squalestatin S和fusarin等骨架上。类似地,对红树林内生真菌Fusarium multiceps的分析鉴定出33个BGCs,其中23个与已知途径缺乏同源性,突显了这些共生体中巨大的隐存化学库。
除了检测之外,计算框架越来越能够推断结构和功能。对超过1000个真菌基因组的比较分析已将数千个预测的BGCs组织成基因簇家族,创建了一个用于促进去重复的注释图谱。此外,在BGC-代谢物对上训练的机器学习模型现在可以从序列数据中以有希望的准确性预测抗菌或细胞毒性活性。在药用植物研究中,这种靶向挖掘已成功地将金丝桃属植物内生菌中的非还原型聚酮簇定位为金丝桃素样抗癌代谢物的假定来源。
内生菌中次生代谢产物的生物合成受多层次等级结构控制,涉及通路特异性转录因子、Velvet家族等全局调控因子,以及表观遗传染色质重塑。由于许多BGCs在标准无菌条件下保持沉默,因此需要先进的分子工具包来释放其潜力。CRISPR-Cas技术,特别是转录激活(CRISPRa)和精确基因组编辑,提供了唤醒这些沉默基因簇的稳健方法。通过删除全局负调控因子、替换天然启动子或修饰表观遗传景观,研究人员可以系统地触发次生代谢产物产生,并捕获大型BGCs进行异源表达,从而绕过传统培养修饰的限制。这些不同调控层次及其对应的靶向激活策略的系统性总结见表2。
内生菌研究中的一个基本瓶颈是无菌培养中营养丰富、均质的条件与宿主组织中资源有限、化学复杂的环境之间的差异。标准培养基通常有利于快速生长的机会菌,无法回收代谢特化的内生菌,也无法诱导表达宿主依赖性BGCs。
模拟内生层。 最近的“环境模拟”策略试图弥合这一差距。基于叶片的培养垫(通过在加工过的叶片组织上覆盖琼脂制成)与标准R2A培养基相比,显著提高了难培养微生物群的回收率。类似地,根据发芽大豆营养概况定制的种子模拟培养基揭示了与盐胁迫相关的区室特异性功能性状。然而,这些离体系统仍然缺乏主动运输和动态免疫反应。
体外-活体植物差距。 功能差异仍然是一个挑战;双重培养筛选常常产生“假阳性”——那些在体外抑制病原体但因生态限制而无法在植物体内定植或保护宿主的菌株。紫锥菊中的感染模型进一步强调,定植结果严格受宿主基因型和组织身份支配。展望未来,真正具有预测性的平台将需要器官型培养和模拟自然内生层时空梯度和信号复杂性的微流控“根-芯片”设备。
预测植物-内生菌系统的行为需要融合生态环境与宿主-微生物基因型的模型。基于约束的代谢建模,特别是基因组规模模型(GEMs),通过将基因组数据嵌入化学计量框架,起着关键的桥梁作用。例如,使用细菌GEMs的模拟已成功预测了叶际群落组装,准确率超过89%,证明了代谢交叉喂养如何构建微生物生态位。
未来的统一模型正朝着系统性的多组学整合发展。通过将植物多尺度模型(从基因组到表型组)与微生物GEMs耦合,研究人员可以将基因调控网络与整个植物的生理学和生态结果联系起来。实现这一愿景取决于将机制重建与机器学习相结合,并遵守FAIR数据标准,以确保稳健、迭代的模型改进。
2.6. 通过联合组学和代谢组学将BGCs与代谢物关联起来
将孤儿BGCs与其产物联系起来是天然产物发现中的主要瓶颈。基于相关性的代谢基因组学已成为领先的解决方案,通过统计分析大型菌株集合中BGCs与代谢物的共现模式,成功地将它们配对。
互补策略利用化学相似性指标和机器学习,即使在较小数据集中也能预测BGC-代谢物关联。验证依赖于稳健的异源表达平台或条件依赖性多组学,这些已成功阐明了数十种隐存途径。这个整合流程——从基因组挖掘到实验验证——对于解码内生层中复杂的化学服务至关重要。
更多结果和补充图表:doi:10.3390/metabo16030164
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!