神经元如何快速重塑表面蛋白以构建精密回路?本研究开创“内吞组学”技术,将蛋白质组学与单细胞转录组学、多组学整合相结合,首次在果蝇嗅觉神经元中系统绘制了发育期轴突的“表面组”与“内吞组”图谱,涵盖超过1000种蛋白。研究发现,内吞作用动态调控细胞连接蛋白的亚细胞定位:Crb蛋白在内体与表面间的精准平衡,是发育期轴突修剪与成熟期轴突完整性的关键开关。进一步多组学整合揭示,邻近神经元分泌的Pvf3因子通过调控其受体Pvr的内吞,指导轴突精准靶向。内吞组学为解析神经发育与疾病的表面蛋白动态提供了全新平台。
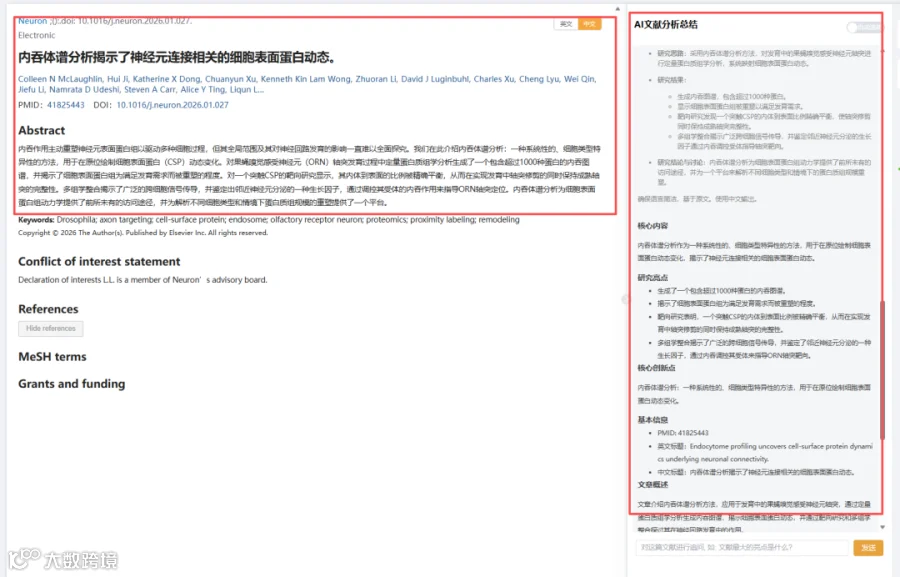
今天给大家解读一篇3月发表在《Neuron》上的题目为“Endocytome profiling uncovers cell-surface protein dynamics underlying neuronal connectivity.”的文章。文章介绍内吞体谱分析方法,应用于发育中的果蝇嗅觉感受神经元轴突,通过定量蛋白质组学分析生成内吞图谱,揭示细胞表面蛋白动态,并通过靶向研究和多组学整合探讨其在神经回路发育中的作用。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!
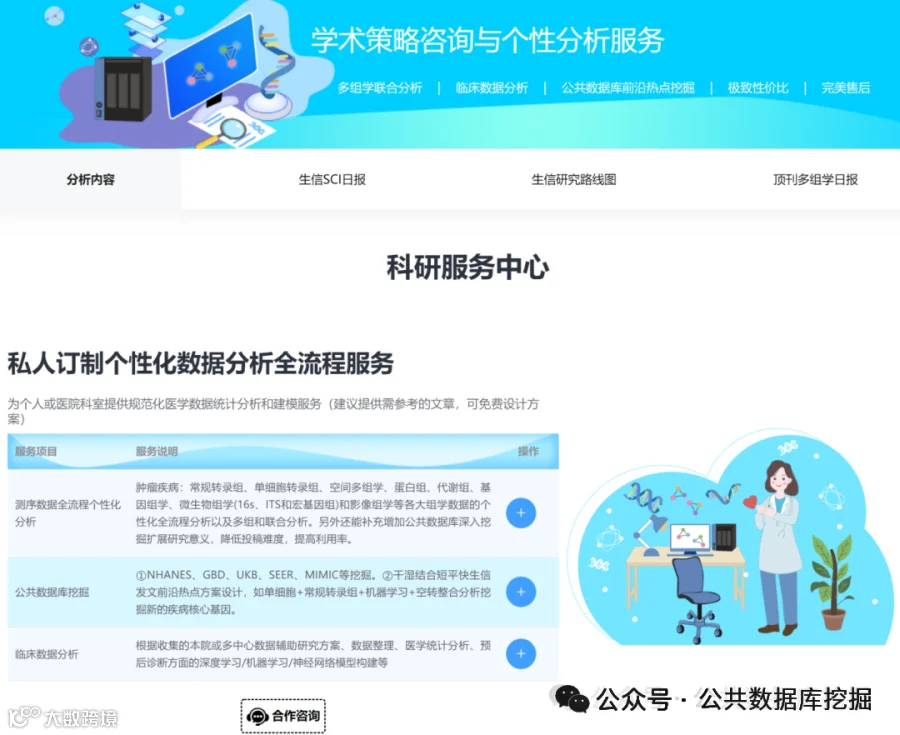
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!


团队成员合影(位于上海陆家嘴中心,可随时预约参观)
题目:《内吞体谱分析揭示了神经元连接相关的细胞表面蛋白动态》Endocytome profiling uncovers cell-surface protein dynamics underlying neuronal connectivity
发表期刊:Neuron
影响因子:15
研究背景:
内吞作用主动重塑神经元表面蛋白组以驱动多种细胞过程,但其全局范围及其对神经回路发育的影响一直难以全面探究。
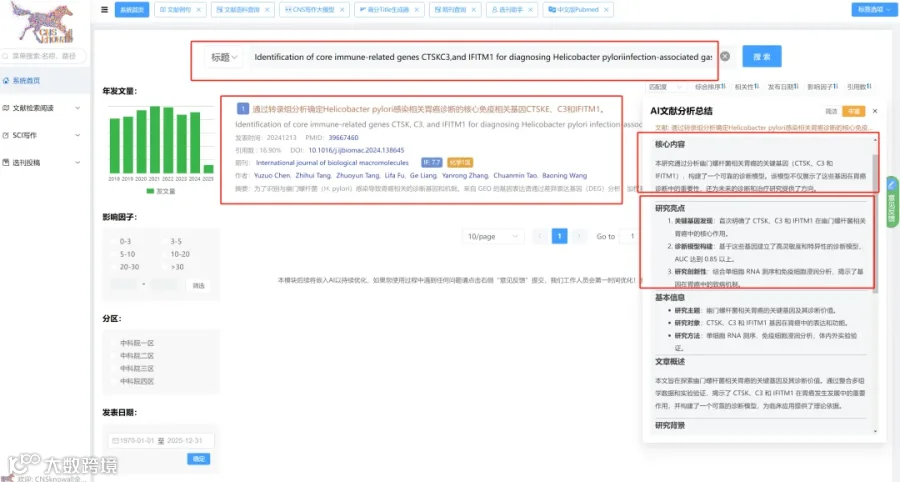
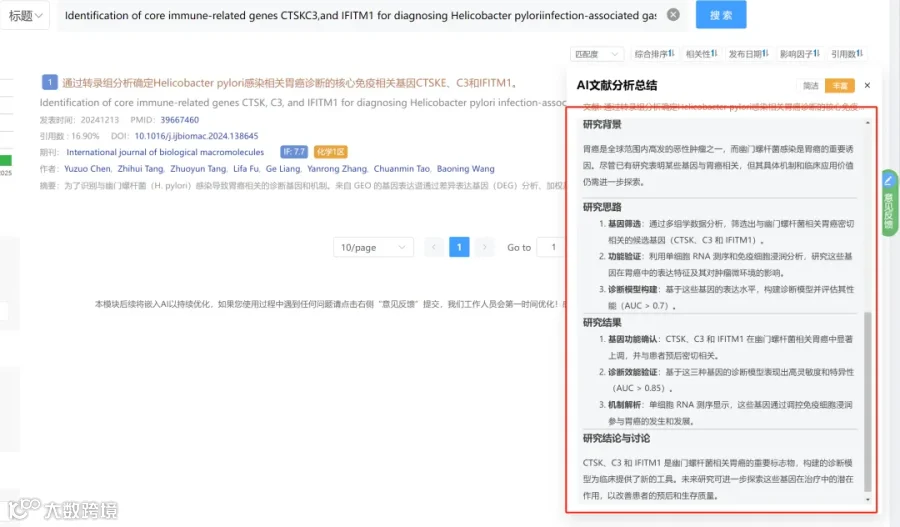
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
采用内吞体谱分析方法,对发育中的果蝇嗅觉感受神经元轴突进行定量蛋白质组学分析,系统映射细胞表面蛋白动态。
研究亮点:
-
-
揭示了细胞表面蛋白组为满足发育需求而被重塑的程度。
-
靶向研究表明,一个突触CSP的内体到表面比例被精确平衡,从而在实现发育中轴突修剪的同时保持成熟轴突的完整性。
-
多组学整合揭示了广泛的跨细胞信号传导,并鉴定了邻近神经元分泌的一种生长因子,通过内吞调控其受体来指导ORN轴突靶向。
研究结果:
-
定量蛋白质组学分析生成了一个内吞图谱,包含超过1000种蛋白。
-
揭示了细胞表面蛋白组为满足发育需求而被重塑的程度。
-
对一个突触CSP的靶向研究表明,其内体到表面的比例被精确平衡,从而在实现发育中轴突修剪的同时保持成熟轴突的完整性。
-
多组学整合揭示了广泛的跨细胞信号传导,并鉴定了邻近神经元分泌的一种生长因子,通过内吞调控其受体来指导ORN轴突靶向。
研究总结:
内吞体谱分析为细胞表面蛋白组动力学提供了前所未有的访问途径,并为一个平台来解析不同细胞类型和情境下的蛋白质组规模重塑。
结果译文:
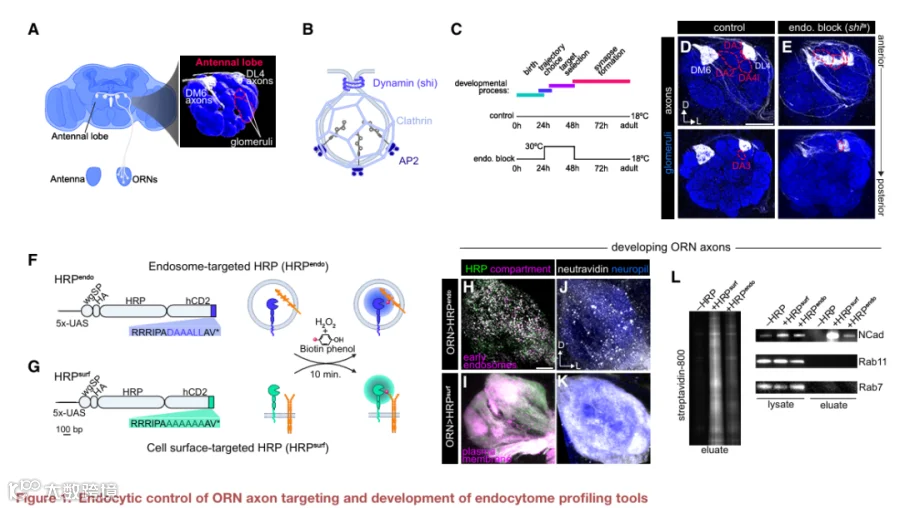
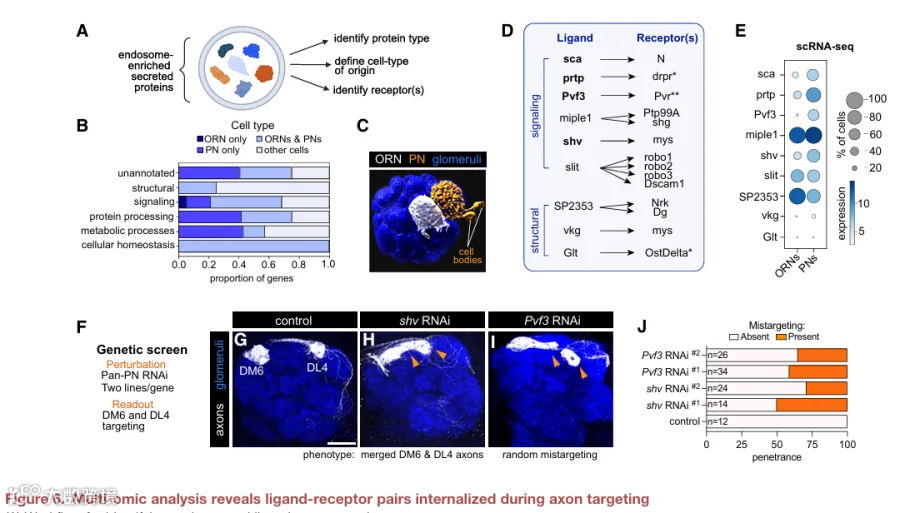
已知CSP内吞作用影响轴突导向,但其在后续靶标选择步骤(即神经元从众多突触后伙伴中选择特定伙伴)中的作用尚不清楚。我们利用果蝇嗅觉受体神经元对此进行了研究。ORN是嗅觉系统的主要感觉神经元。进入大脑后,约50种不同ORN亚型的轴突遵循特定轨迹,靶向其突触后伙伴(约50种投射神经元亚型)的树突,在称为小球的结构单元内形成一对一的突触连接(图1A)。
网格蛋白介导的内吞作用是神经元中主要的受体内化途径。它涉及衔接蛋白(如AP2)的顺序招募以聚集内吞货物,随后招募网格蛋白和GTPase发动蛋白,后者介导囊泡从质膜上断裂(图1B)。由于早期的轴突导向决策(如轨迹选择)会影响靶标选择,我们通过仅在DM6和DL4小球投射的ORN轴突中表达温度敏感的shibire转基因(shits,编码发动蛋白的显性负性形式),将内吞抑制在时间上限制于后一过程(图1C-1E)。
在大多数轴突完成轨迹选择之后、但在靶标选择和突触组装之前,将这些果蝇在30°C下饲养24小时,导致ORN靶向出现稳健且特异的缺陷(图1C-1E)。当发动蛋白活性被阻断时,DM6-ORN轴突向它们正常小球的靶向显著减少,而向两个特定小球(DA2和DA4l)的神经支配显著增加(图1E和S1A)。同样,损害AP2或网格蛋白的表达或功能也会导致DM6-ORN轴突出现刻板的错误靶向(图S1C和S1D)。同样,阻断DL4-ORN中的发动蛋白活性导致它们错误靶向DA3小球(图1E和S1B)。在特定发育窗口期损害内吞作用导致ORN轴突支配刻板但错误的小球,这一事实表明该过程调节了轴突靶标选择所必需的特定CSP库。
为了全面定义轴突靶向期间内体中富集的CSP,我们开发了一种定量邻近标记方法,以绘制发育中ORN轴突中CSP的表面-内体分布。CSP通过其胞质尾部的短氨基酸基序被选择进行网格蛋白介导的内吞。因此,我们将邻近标记酶辣根过氧化物酶靶向到内体腔中(以下称为HRPendo),方法是将HRP置于人CD2(一种单次跨膜蛋白)的N端,并在其胞质尾部添加一个基于双亮氨酸的内吞基序(图1F)。整个开放阅读框处于UAS控制之下,用于GAL4介导的细胞类型特异性表达(图1F)。我们验证了HRPendo在体外和体内均被内化(图S2A和S2C),并且与内溶酶体系统的标志物共定位(图S2B和S2D-S2F)。此外,在发育中的ORN轴突中,HRPendo与mCherry-2xFYVE部分共定位,后者主要积聚在早期内体上(图1H),验证了它在我们的目标细胞中处于正确的亚细胞区室。这些数据共同表明,网格蛋白介导的内吞为将HRP递送到内体提供了一条有效途径。
我们将HRPendo的双亮氨酸基序突变为丙氨酸,从而标记驻留在质膜上的CSP,以下称为HRPsurf(图1G)。HRPsurf应与HRPendo一样通过分泌途径到达质膜,但不会稳健地进入内体。我们验证了HRPsurf在ORN轴突和培养细胞中主要定位于质膜(图1I、S2A和S2B)。
在施加H2O2后,HRPendo和HRPsurf均使用膜通透性的生物素-酚在解剖的大脑中标记蛋白质(图1J和1K)。生物素分布反映了它们各自的亚细胞区室,HRPendo标记呈点状,而HRPsurf在ORN轴突中呈连续状(图1J和1K)。链霉亲和素磁珠富集证实了多种蛋白质的标记(图1L,左),免疫印迹验证了区室特异性——在链霉亲和素磁珠洗脱液中检测到神经元表面蛋白N-钙粘蛋白,但未检测到胞质内体相关Rab GTP酶Rab7和Rab11(图1L,右)。这些结果与HRP朝向胞外和腔隙空间的取向以及其产生的生物素-苯氧基自由基的膜不通透性一致,后者阻止了标记胞质驻留蛋白(如Rabs)。
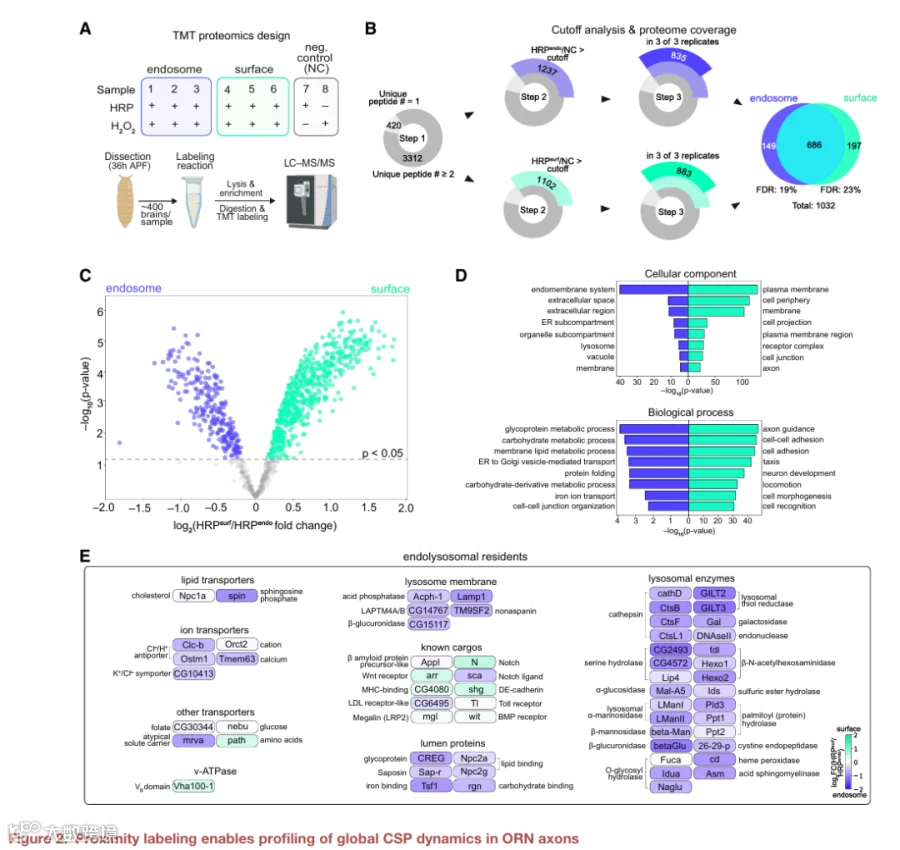
接下来,我们将HRP介导的生物素化与基于质谱的蛋白质组学相结合,以定义ORN轴突的“内吞组”和“表面组”。我们专门在靶标选择期间对ORN轴突进行谱分析,因为我们可以轻松地将远端轴突与其在触角中的胞体分离(图1A)。使用基于串联质量标签的定量策略,我们分析了HRPendo或HRPsurf的三个生物学重复,以及两个阴性对照(缺乏HRP或H2O2),以排除非特异性标记(图2A)。
从质谱检测到的总共3,732种蛋白质中,我们通过四个过滤步骤鉴定了区室富集的蛋白质。首先,我们保留了检测到至少两条独特肽段的3,312种蛋白质(图2B;表S2)。其次,我们按实验组与阴性对照的TMT比值降序排列蛋白质,并验证了含有信号肽和/或跨膜结构域的蛋白质富集,而缺乏这些特征的假定污染物则没有富集(图S3A-S3D)。按照已发表的方案,我们使用这种比值法过滤掉污染物(图2B)。第三,由于HRPendo或HRPsurf重复样本高度相似(图S3E),我们保留了每个条件下所有三个蛋白质组中均存在的蛋白质(图2B)。这些步骤共得到1,032种蛋白质,其中大多数被HRPendo和HRPsurf标记同时检测到(图2B),突出了蛋白质定位的动态性质。最后,为了确定区室特异性富集,我们比较了这些蛋白质在HRPendo与HRPsurf标记蛋白质组中的相对丰度。这分别得到了248种和571种在内吞组和表面组中显著富集的蛋白质(图2C)。
证实了我们方法的空间特异性,基因本体论分析将内吞组蛋白分类到胞外区域、内膜系统和溶酶体,而表面组蛋白则定位到质膜和细胞外围(图2D)。与所分析的阶段一致,表面组中排名靠前的生物学过程GO术语与轴突发育和细胞粘附相关(图2D)。值得注意的是,细胞-细胞连接组织术语在内吞组中高度富集(图2D),表明这类特定蛋白质的内化对于轴突发育很重要(见图4和图5)。内吞组富集了代谢和离子转运术语(图2D),与其在大分子降解中的作用一致。对这些类别蛋白质的深入研究发现,其中包含溶酶体酶、整合膜蛋白和膜转运蛋白(图2E),建立了内溶酶体驻留蛋白的综合图谱,并进一步验证了我们的谱分析。
我们的内体谱分析还捕获了分泌途径与内溶酶体网络之间的串扰,证据是高尔基体/内质网驻留蛋白的存在以及跨高尔基体网络的驻留蛋白(图2E),这些蛋白参与内体与这些细胞器之间的物质运输。此外,我们发现内吞组富集了参与选择性自噬的蛋白质(图2E),选择性自噬是一种通过自噬体将错误折叠的蛋白质递送到溶酶体进行降解的途径,以及参与内质网到溶酶体相关降解的蛋白质,这是一种最近描述的在应激期间清除蛋白质聚集体和错误折叠蛋白质的途径。这些发现强调了我们的方法捕捉与内体区室密切相关的细胞过程的能力。
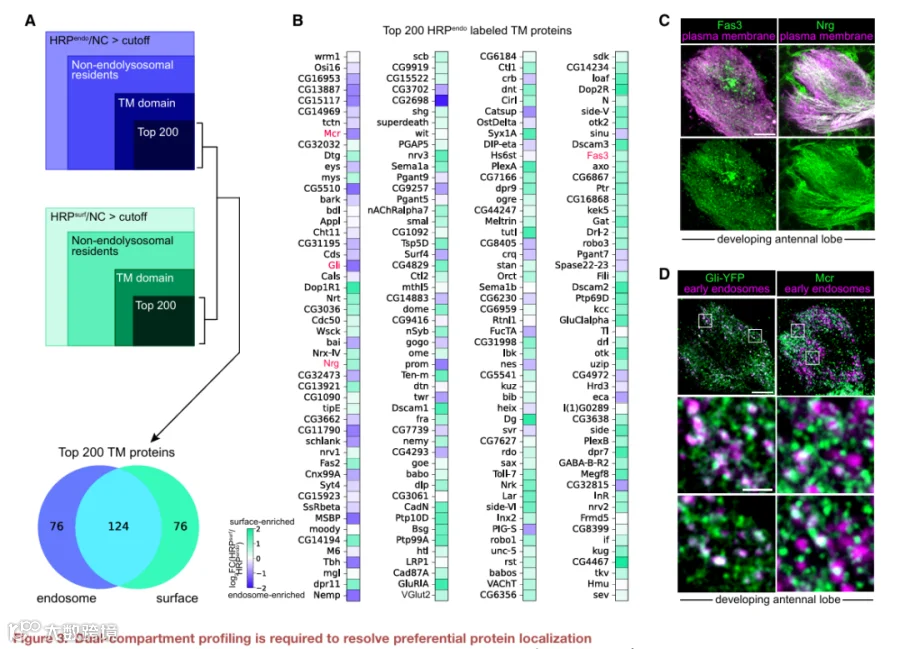
4.内吞组含有多个对轴突发育至关重要的细胞-细胞连接蛋白
我们首先关注内体中最富集的CSP货物。与表面组中检测到许多典型的神经发育蛋白(图S4A和S4B)相反,细胞-细胞连接CSP在内吞组中高度富集(图2D)。这些连接包括隔膜连接、紧密连接和粘附连接,由将细胞彼此连接或连接到细胞外基质的CSP形成。在神经系统中,此类连接在祖细胞、胶质细胞、髓鞘和轴突之间形成,但不在神经元之间形成。由于ORN轴突在靶标选择期间与胶质细胞接触极少,这些连接蛋白在内体中的富集是出乎意料的。
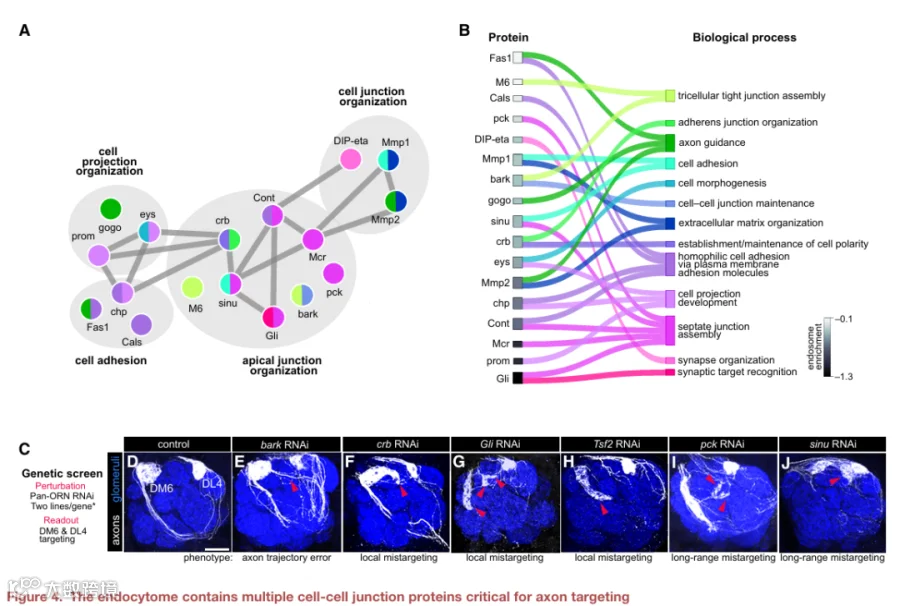
这促使我们对细胞-细胞连接、细胞粘附和细胞投射组织类别中的蛋白质进行聚焦分析,因为其中许多蛋白质存在相互作用(图4A)。我们为每种蛋白注释了其最可能涉及的特定生物学过程。少数蛋白质已知在神经元中发挥作用,但许多则不然(图4B)。我们先前的共定位实验证实了两种连接蛋白——巨球蛋白补体相关蛋白和Gli在内体中的富集(图3D)。
接下来,我们进行了靶向功能缺失筛选,以评估这些连接蛋白是否在轴突发育中发挥作用。我们在所有ORN中表达针对每个基因的RNAi,并监测投射到DM6小球的轴突(图4C)。所有测试的六个基因均产生轴突表型(图4D-4J;表S4)。例如,清道夫受体bark beetle促进轴突轨迹选择,但对后期发育事件影响不大(图4E)。缺失粘附连接蛋白crumbs或隔膜连接成分Transferrin 2和Gli导致轴突错误靶向到DM6附近的小球(局部错误靶向;图4F-4H),而敲低claudins pickel和sinuous则导致长程错误靶向(图4I和4J)。这些数据表明,连接CSP控制着轴突靶向的不同方面。最近一项研究发现,紧密连接和粘附连接CSP的转录本在哺乳动物神经元神经突生长和突触发生期间富集,表明这类蛋白可能是回路组装的保守调节因子。
由于这是对ORN表面组的首次谱分析,我们还检测了表面富集的CSP是否调节轴突发育。我们测试的6个基因中有5个控制着轴突靶向的一个或多个方面(图S4C-S4I;表S4)。测试的CSP中有91%(11个中的10个)产生表型,这既验证了我们谱分析的生物学相关性,也验证了回路组装依赖于分布在内体和质膜之间的多个CSP的观点。
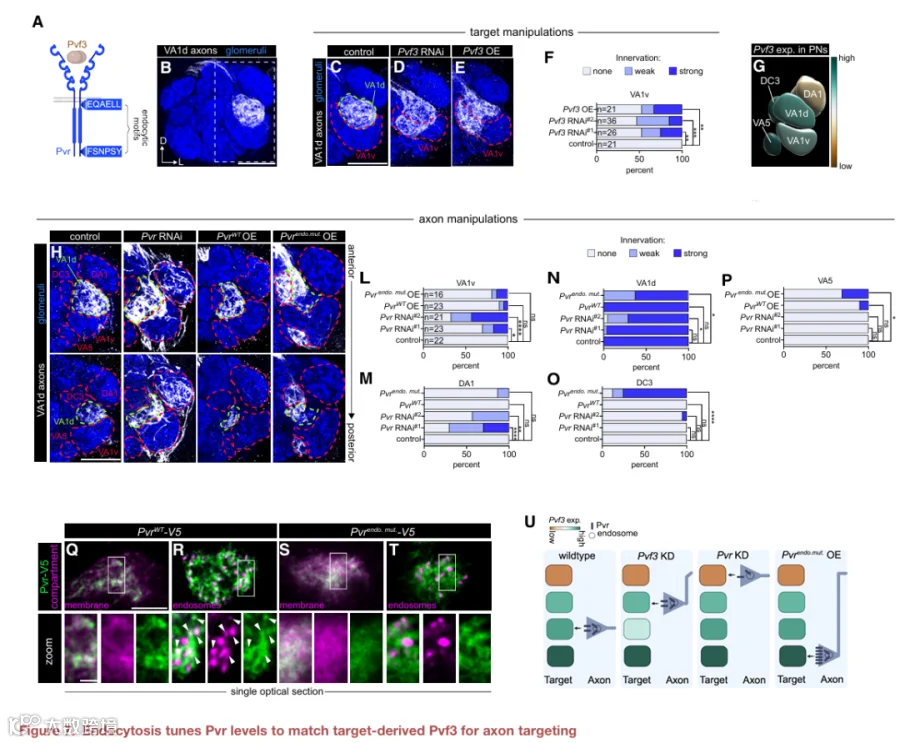
5.Crumbs促进轴突修剪并通过内吞作用保护轴突完整性
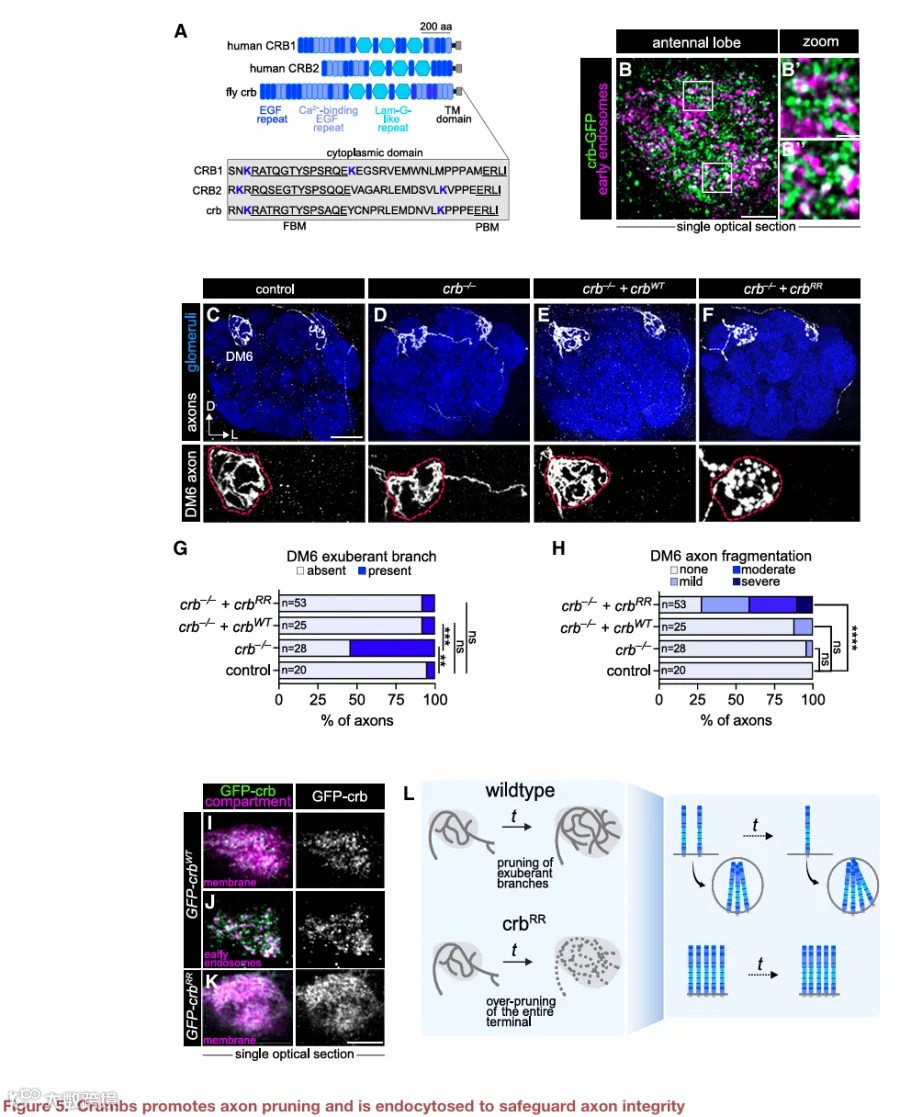
内吞作用如何塑造连接CSP的功能?我们通过研究crb来探讨这个问题,crb是一种进化上保守的蛋白质,参与上皮细胞极性、粘附连接形成和光感受器完整性。人类有三种CRB蛋白,其中CRB1和CRB2与单个果蝇crb最相似(图5A,顶部)。尽管CRB1/2在大脑中表达,但任何crb蛋白在该组织中的神经元作用很大程度上尚未被定义。
根据我们的发育期单细胞RNA测序数据,crb在ORN中表达,但在其突触后伙伴PN中不表达(图S5A),表明我们内吞组中的crb很可能是由ORN自主产生的。使用内源标记的crb等位基因,我们发现crb-GFP蛋白呈点状,并与发育中ORN轴突中的早期内体部分共定位(图5B’和5B”),非共定位的点状结构可能位于其他内体区室中。
在验证了crb作为内体富集蛋白的蛋白质组学鉴定后,我们接下来研究了它在轴突发育中的功能。使用可抑制细胞标志物的嵌合分析,我们利用无效等位基因探究了crb的细胞自主作用。我们使用eyeless-FLP或热激-FLP分别诱导大克隆(占ORN的30%-50%)或小克隆(低至单个ORN),其中只有纯合crb突变轴突在DM6-和DL4-ORN轴突中表达膜标记。与对照组相比,突变克隆显示出明显更多的超出DM6小球的过度分支(图5C、5D、5G、S5B、S5C和S5F)。通过仅在标记的ORN中表达野生型crb,可以挽救这种表型(图5E、5G、S5D和S5F),验证了crb以细胞自主方式发挥作用。在发育过程中,DM6轴突会向邻近小球发出分支,这些分支随后被修剪以形成成年回路。它们在crb突变体中的持续存在表明crb促进了过度分支的修剪。
内吞作用是否在crb依赖的修剪中发挥作用?Crb高度保守的37个氨基酸胞质尾部包含两个赖氨酸残基(图5A),其泛素化驱动内吞。我们比较了野生型crb挽救转基因与内吞缺陷版本(UAS-GFP-crbRR)产生的crb蛋白的分布,后者除两个胞质赖氨酸突变为精氨酸外,序列、基因组插入位点和蛋白表达水平均相同(图S5M)。CrbWT呈点状并与早期内体标志物部分共定位(图5I和5J)。CrbRR均匀分布在膜上(图5K),并且在早期内体区室中基本不存在(图5L),与其内化受损一致。
在crb突变ORN中表达crbRR并没有改变成年DM6轴突中存在的过度分支数量(图5F、5G、S5E和S5F),表明crb内吞作用并非轴突修剪所必需。此外,发育中的crbRR轴突向附近小球延伸的分支比例与对照组相似(图S5K和S5L),证明将crb限制在质膜上并不会损害轴突发育的正常特征。总之,这些结果表明,是质膜上的crb(而非内体中的crb)对于去除过度分支是必需的。
由于我们的数据暗示crb促进发育性轴突修剪,我们询问持续的crb表面定位是否会导致成熟轴突过度修剪。引人注目的是,抑制crb内吞导致轴突断裂(图5F,底部,5H,S5E和S5G)——轴突不稳定的标志。与对照组相比,crbRR中ORN细胞体的数量没有变化(图S5N),表明这种表型不是细胞死亡的结果。因此,crb的长期表面定位导致整个轴突末端的过度修剪。我们提出,crb在内体与细胞表面之间的精确平衡,使得在发育过程中能够进行轴突修剪,同时保留成熟回路中的末端完整性(图5M)。总之,这些发现凸显了空间解析的CSP谱分析如何揭示协调发育回路精修和长期回路稳定性的关键机制。
内吞组还包含多种分泌蛋白。由于这些蛋白可能来自邻近的神经元或胶质细胞,并通过受体介导的内化进入ORN,我们询问是否可以进一步利用该数据集来推断细胞-细胞相互作用(图6A)。
84种内体富集的分泌蛋白(表S2)跨越五个功能类别:(1)细胞稳态,(2)蛋白加工,(3)信号传导,(4)代谢过程,以及(5)结构作用(图6B),突显了轴突正在导航的丰富细胞外环境。信号传导蛋白是最大的类别,既包含经典的神经发育调节因子(例如slit和scabrous),也包含先前未与发育关联的蛋白质(表S2)。为了开始解码轴突发育背后的细胞-细胞通讯,我们询问哪些细胞类型分泌了ORN内化的蛋白。利用我们的发育期scRNA-seq数据,我们将每种蛋白的表达映射到ORN或PN(图6C)。许多转录本在ORN和PN之间共享(图6B和S6)。尽管只有两种转录本在ORN中高表达,但超过35%的未注释、蛋白加工和代谢过程类别中的转录本,以及15%的信号传导类别中的转录本,在PN中表达,但在ORN中不表达(图6C),表明内吞组包含来自邻近神经元而非ORN自身的分泌蛋白。为了测试这一点,我们关注了血小板衍生生长因子和血管内皮生长因子相关因子3,这是一种已知在血细胞迁移和存活中起作用的分泌性生长因子。Pvf3在PN中高度富集,但在ORN中则不然(图6C和S6B),使其成为探究跨细胞信号传导在ORN轴突靶向中作用的理想候选者。
Pvf3与其受体Pvr(一种受体酪氨酸激酶)结合。Pvr的C末端尾部包含多个内吞基序(图7A),我们推测Pvr可能通过内吞作用被调节。与这一想法一致,Pvr在ORN轴突的内吞组中富集(图S7),并且在发育中的ORN轴突中与早期内体共定位(图S8A)。利用分裂GAL4系统,我们证实了Pvf3在多种PN亚型中表达,包括VA1d-PN(图7G、S8B和S8C),其中VA1d-ORN轴突进行靶向(图7B)。PN转录组中Pvf3表达的显著异质性(图S8C)使我们假设细胞类型特异性的配体水平指导ORN轴突靶向。
由于在发育过程中可以同时操作VA1d-PN和VA1d-ORN,我们得以研究PN来源的Pvf3在调节ORN轴突靶向VA1d小球中的作用(图7B)。Pvf3在VA1d-PN中表达水平相对较高(图7G和S8C)。在VA1d-PN中敲低Pvf3导致VA1d-ORN错误靶向到邻近的VA1v小球(图7C、7D和7F)。相反,在PN中过表达Pvf3导致VA1d-ORN轴突过度生长,超出VA1d小球边界并延伸到相邻的VA1v小球(图7E和7F)。这些数据表明,Pvf3作为指导信号,促进ORN轴突靶向其正确的小球。
Pvr在ORN中表达。我们假设Pvr的内吞调节其表面水平,以匹配来自PN的Pvf3信号。为了验证这一点,我们在ORN中表达野生型Pvr或内吞缺陷型Pvr突变体(图7A)。ORN中Pvr的敲低导致VA1d-ORN轴突的错误靶向(图7H、7L和7M),这可以通过表达野生型Pvr来挽救(图7I、7M和7N)。引人注目的是,表达内吞缺陷型Pvr(Pvrendo.mut.)导致VA1d-ORN轴突异常,类似于Pvf3过表达的表型——轴突过度生长超出VA1d小球边界(图7K和7P),而野生型Pvr没有这种效应(图7J、7O和7P)。相比之下,当在VA1d-ORN中表达PvrWT或Pvrendo.mut.时,相邻小球(VA1v、DC3、DA1和VA5)中ORN轴突的神经支配没有差异(图7L-7P)。在培养的S2细胞中,PvrWT-V5与早期和晚期内体标志物共定位,而Pvrendo.mut.-V5在细胞中呈均匀分布(图S9)。在发育中的ORN轴突中,PvrWT-V5呈点状并与早期内体标志物部分共定位,而Pvrendo.mut.-V5则均匀分布在质膜上(图7Q-7T)。这些数据共同表明,Pvr的内吞对于微调ORN轴突中Pvr的水平以响应PN来源的Pvf3信号至关重要,从而指导精确的轴突靶向(图7U)。
更多结果和补充图表:doi:10.1016/j.neuron.2026.01.027
长按二维码关注我们,用最短的时间和最高的效率学习更多数据分析方法!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:
https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的科研数据(0代码生信+统计学)分析平台,同时含有机制图模块+汉化版Pubmed融合Deepseek高效筛选目标文献+SCI文献例句/语料检索模块+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动批阅!