今天给大家解读一篇4月发表在《Microorganisms》上的题目为“Environmental Regulation of Gut Microbial Networks Links to Growth Variation in Schizopygopsis younghusbandi Across Contrasting Aquaculture Systems.”的文章。该研究旨在探究不同养殖水环境如何通过影响肠道微生物网络进而影响拉萨裸裂尻鱼的生长变异。实验选取五种典型养殖系统,饲养3龄幼鱼90天后测定生长指标,并采集水样和肠道样本进行宏基因组测序。结果发现了显著的生长差异、微生物群落结构差异以及环境-微生物关联。循环水养殖系统提供了更稳定的水温、更高的营养调控能力,其水体及肠道微生物网络复杂度最高,鱼生长最佳。湿地系统水温低、网络连接弱,生长和存活最差。研究强调了环境稳定性在塑造有益微生物互作网络、促进宿主生长中的关键作用,并推荐循环水系统作为高原鱼类养殖的优化模式。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!包括数据集、绘图代码、图表复现、思路总结、参考文献……0代码!鼠标点点点即可轻松完成5-10分生信SCI全文复现!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!









团队成员合影(位于上海陆家嘴中心,可随时预约参观)






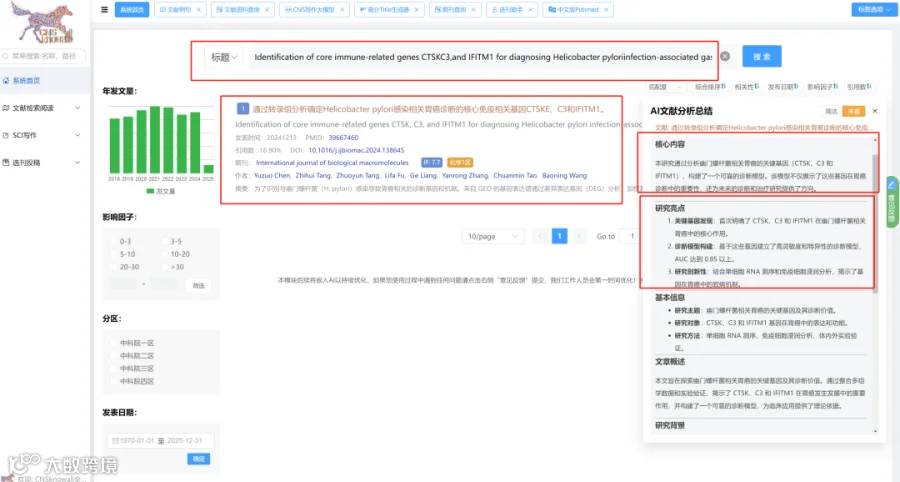
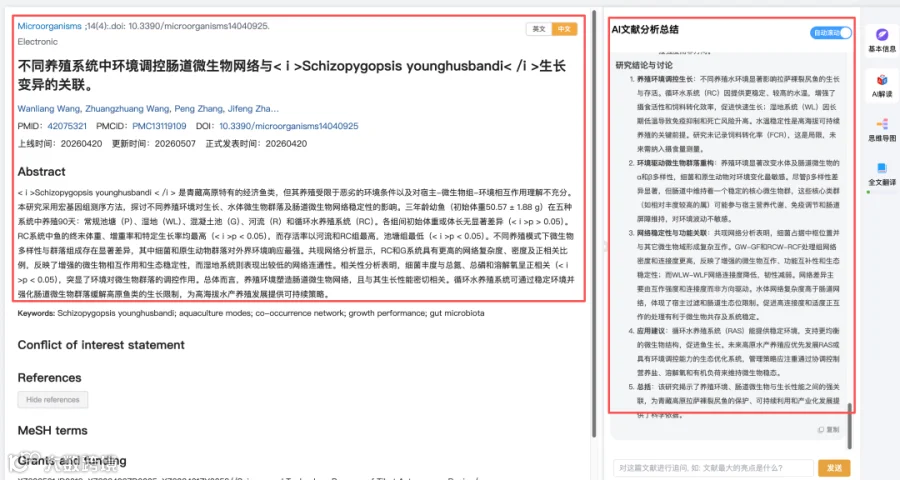
题目:《不同养殖系统中环境调控肠道微生物网络与Schizopygopsis younghusbandi生长变异的关联》Environmental Regulation of Gut Microbial Networks Links to Growth Variation in Schizopygopsis younghusbandi Across Contrasting Aquaculture Systems
发表期刊:Microorganisms
影响因子:4.2
研究背景:
- 产业需求
中国水产养殖发达,但西藏起步晚,高原鱼类生长慢、性成熟迟、养殖周期长,渔业经济与内地差距大。拉萨裸裂尻鱼是青藏高原特有且唯一实现全人工繁殖的裂腹鱼,具有重要经济与生态价值。 - 问题提出
不同养殖水体(池塘、湿地、混凝土池、河流、循环水)具有不同特性,选择合适的养殖模式是提质增效的关键。微生物是水生环境和鱼类肠道的关键组成部分,参与宿主营养吸收、免疫调节、生理稳态。外部环境因素(盐度、营养、养殖条件)可显著重塑肠道微生物结构,但环境-微生物-宿主代谢之间的双向互作机制尚不明确。 - 研究空白
关于拉萨裸裂尻鱼的研究多集中在个体生物学、行为学、分子生物学和遗传进化,尚未有研究报道不同水环境对其健康养殖过程中微生物群落的影响。

研究思路:
- 实验设计
在西藏拉萨和曲水选取5种典型养殖环境(P、WL、G、R、RC),每个环境设3个重复,每个重复放养100尾初始体重(50.57±1.88 g)和体长相近的健康F1代幼鱼。每日投喂两次商品饲料,养殖90天。 - 样本采集与指标测定
-
水质:现场测定水温、溶解氧(DO)、pH,实验室测定BOD5、TN、TP、NH₃-N、NO₂⁻、总硬度、总碱度。 -
生长性能:称重、量体长,计算增重率(WGR)、特定生长率(SGR)、肥满度(CF)、存活率。 -
微生物样本:水样过滤收集,无菌解剖取肠道内容物,液氮速冻后-80℃保存。 - 宏基因组测序与生物信息分析
-
DNA提取、建库、Illumina NovaSeq X Plus测序。 -
质控(fastp)、去宿主(BWA)、组装(MEGAHIT)、基因预测(Prodigal)、非冗余基因集构建(CD-HIT)、物种注释(DIAMOND比对NR数据库)。 - 统计分析
-
单因素方差分析(ANOVA)+ Duncan多重比较检验水质与生长差异。 -
Alpha多样性(Shannon指数)、PCoA分析(Bray-Curtis距离)。 -
随机森林分析鉴定指示物种。 -
Spearman相关与Mantel分析环境-微生物关联。 -
共现网络构建(|R|>0.6, p<0.05)并计算拓扑参数(节点数、边数、网络密度)。 -
RDA分析环境因子对肠道微生物的解释度。
研究亮点:
- 首次系统研究
首次报道了不同养殖水环境对拉萨裸裂尻鱼肠道微生物群落结构的影响。 - 多域微生物整合分析
同时分析了细菌、原生动物、藻类和真菌四个微生物域在水体和肠道中的多样性、组成及互作网络,而非仅关注单一类群。 - 环境-微生物-生长性能关联
通过冗余分析(RDA)、随机森林和共现网络,将环境因子、微生物群落变化与鱼体生长表现直接关联,阐明了环境调控肠道微生物进而影响生长的潜在机制。 - 核心微生物群稳定性
发现尽管不同系统间β多样性显著差异,但肠道内存在一个稳定的核心微生物群,暗示其对宿主基础生理功能的保守性。
研究结果:
- 水质与生长差异
-
水质参数差异显著:RC组水温最高;WL组DO最高;G组TN、总硬度最高。 -
生长表现:初始体重/体长无差异。RC组最终体重、WGR、SGR显著最高(p<0.05),PF组最低。存活率:RF和RCF组最高(与GF无差异),PF组最低。 - 微生物多样性
- 水体
WL组细菌Shannon最高;PW组原生动物、藻类、真菌Shannon最高。 - 肠道
WLF和RF组细菌多样性最高;RCF组原生动物多样性最低;WLF组藻类和真菌多样性最高。 -
PCoA:细菌和原生动物在两种生境中均表现出最强的处理间分离(p<0.001),藻类和真菌分离较弱。 - 微生物组成
- 门水平
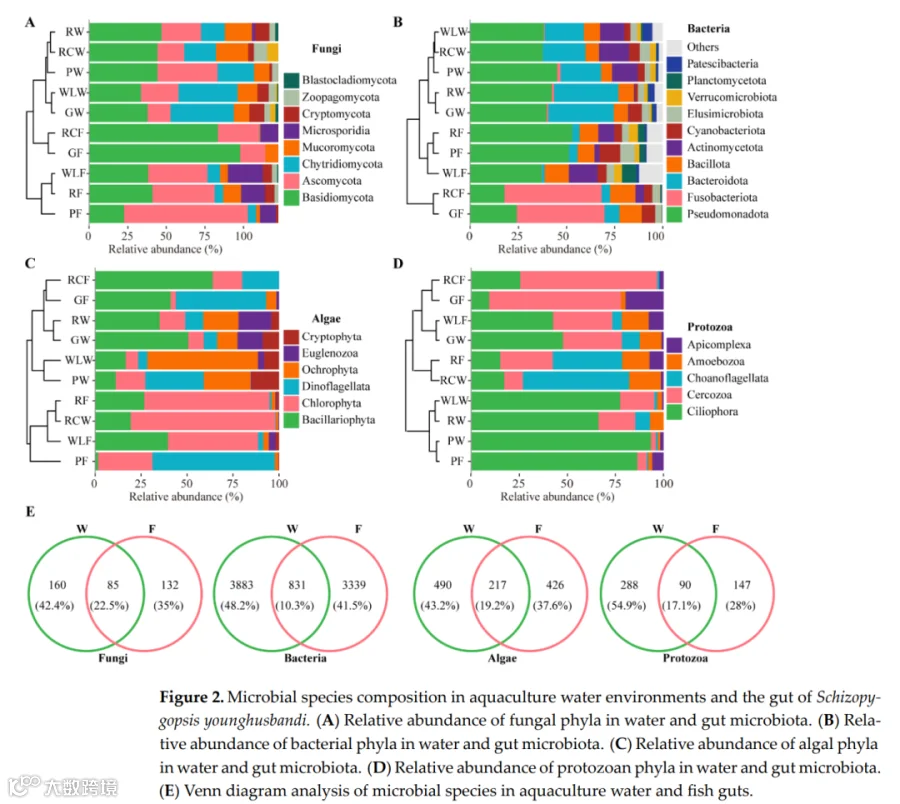
真菌以子囊菌门(Ascomycota)和担子菌门(Basidiomycota)为主;细菌以假单胞菌门(Pseudomonadota)、拟杆菌门(Bacteroidota)和芽孢菌门(Bacillota)为主;藻类以绿藻门(Chlorophyta)和硅藻门(Bacillariophyta)为主;原生动物以纤毛虫门(Ciliophora)和丝足虫门(Cercozoa)为主。 - 共享物种
水体与肠道共享物种比例低(细菌仅10.3%),表明生境过滤和生态位分化强烈。 - 指示物种与环境关联
-
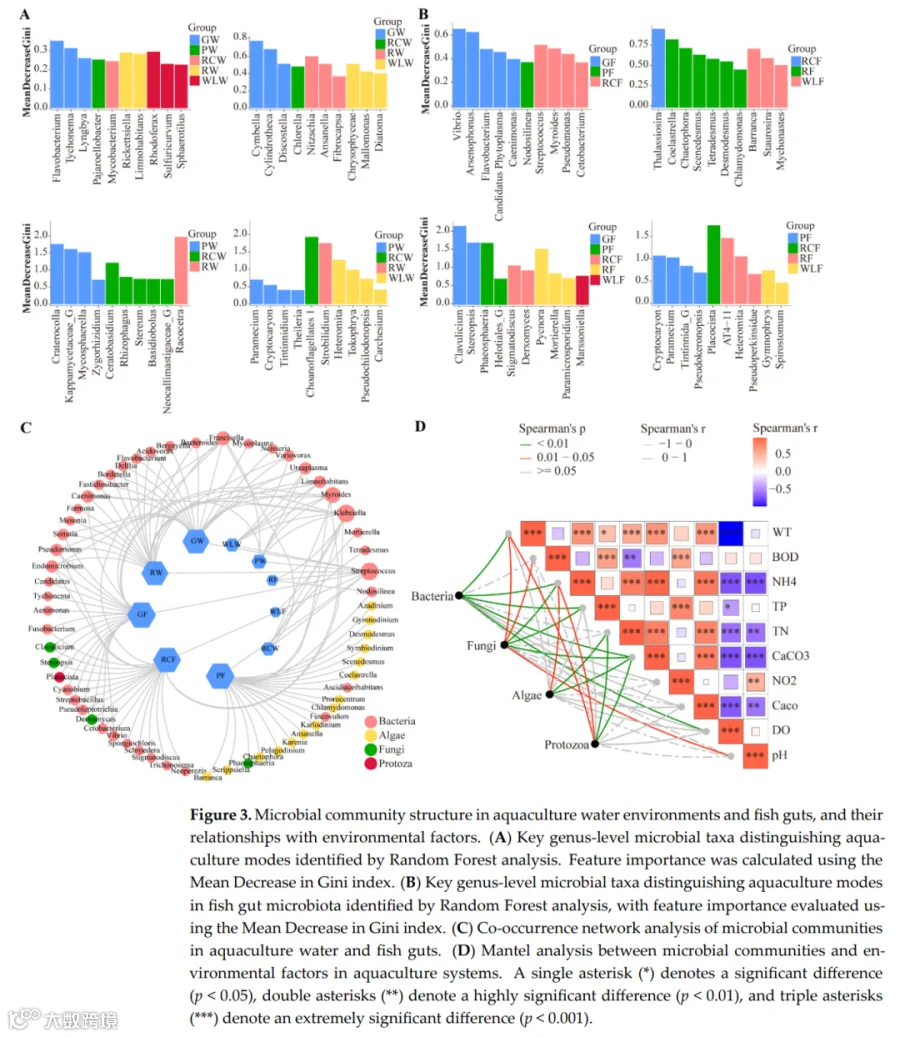
随机森林:水体中,GW组富集黄杆菌属,WLW组富集鞘氨醇单胞菌属,PW组富集湖杆菌属等;肠道中,GF组富集弧菌属,PF和RCF组富集黄杆菌属和假单胞菌属,WLF组富集鞘氨醇单胞菌属和芽孢杆菌属。 -
Spearman/Mantel:细菌丰度与TN、TP、DO正相关(p<0.05);藻类与pH、DO正相关;真菌与NO₂⁻、CaCO₃负相关;原生动物与BOD5、NH₄⁺负相关。 -
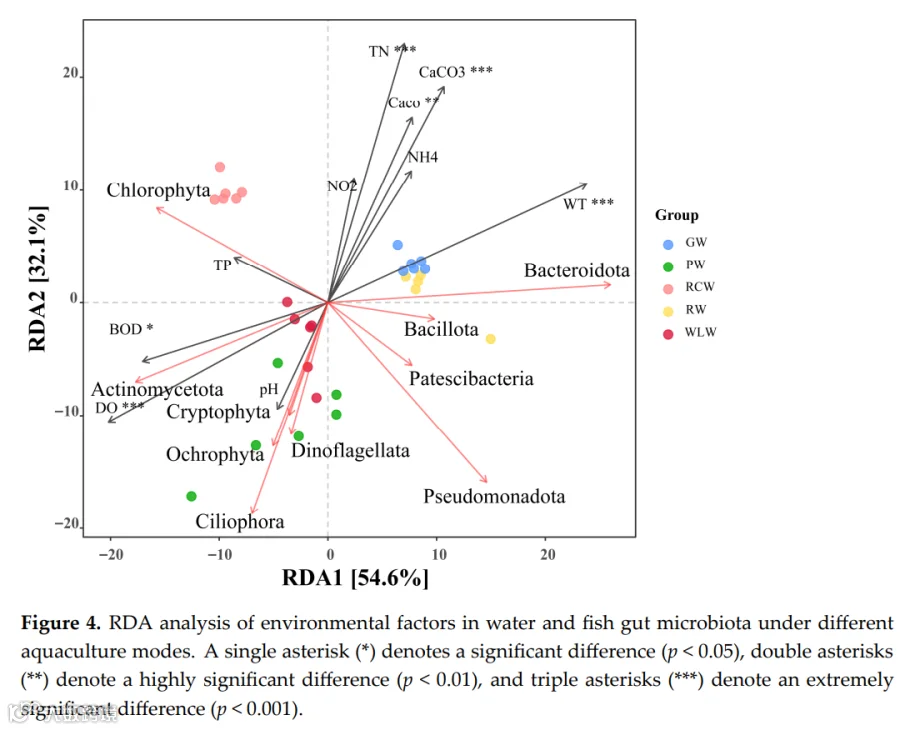
RDA:第一轴解释54.6%,第二轴32.1%,合计86.7%。TN、CaCO₃、CaO、水温是驱动肠道微生物群落变化的主要环境因子(p<0.05)。 - 共现网络
- 水体网络
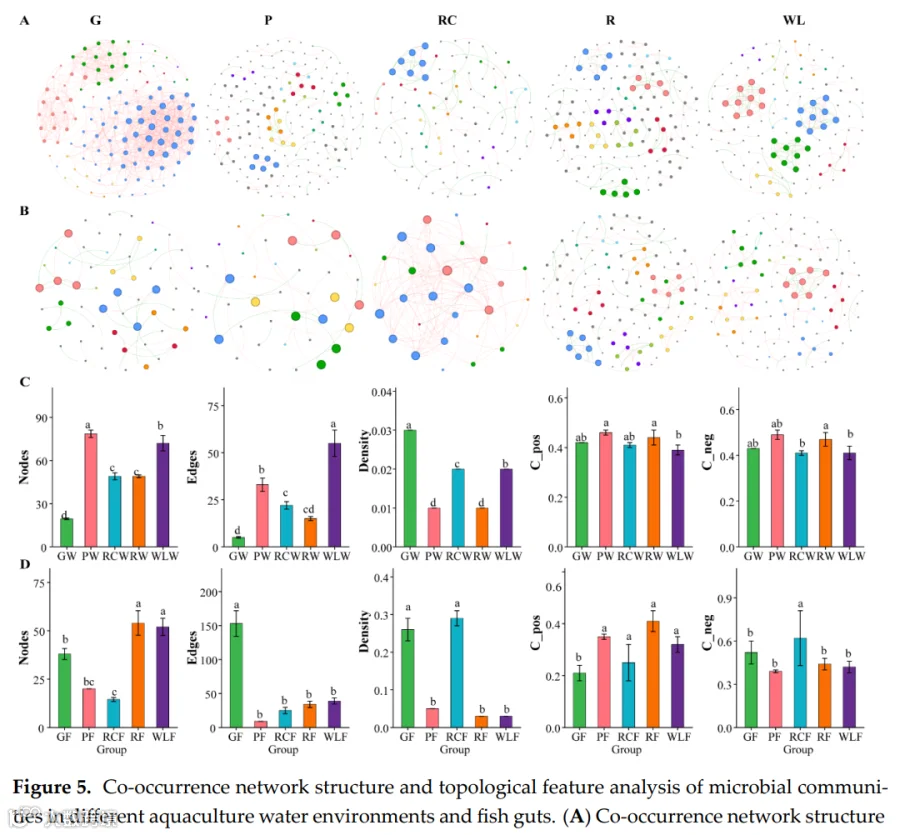
GW和WLW组网络节点数、边数、密度最高(最复杂连接),PW和RCW组最简单。 - 肠道网络
RCF和RF组网络复杂度最高(节点/边/密度显著高于其他组,p<0.05),GF和PF组最低,WLF组中等。 -
网络以细菌为中枢节点,与藻类、真菌、原生动物形成复杂互作。正负相关比例在各组间大致相当,变化主要体现在连接强度而非方向。
研究总结:
结果译文:
1.不同养殖水体的理化参数及其对拉萨裸裂尻鱼生长的影响
2.不同养殖水体及拉萨裸裂尻鱼肠道微生物多样性的变化
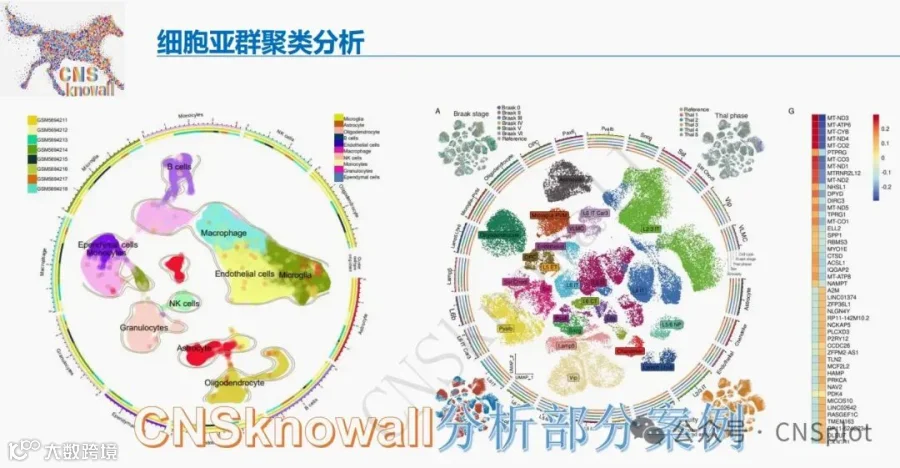
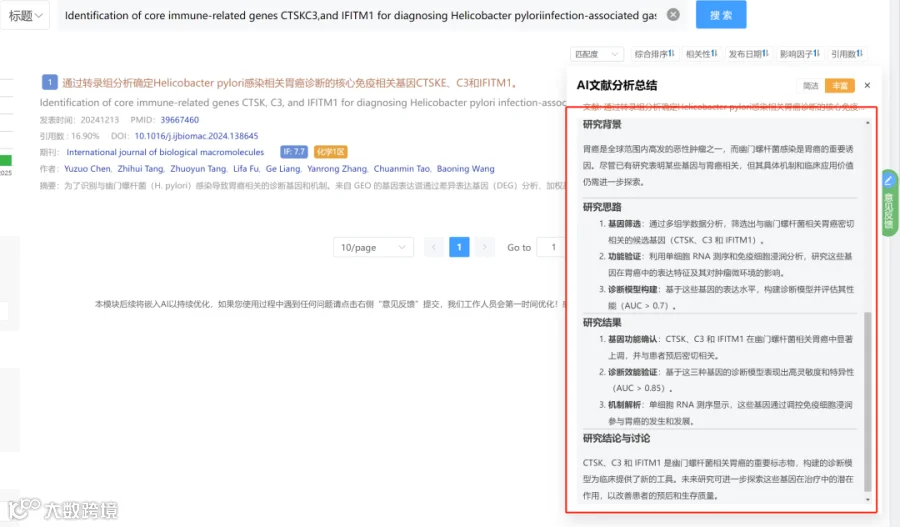
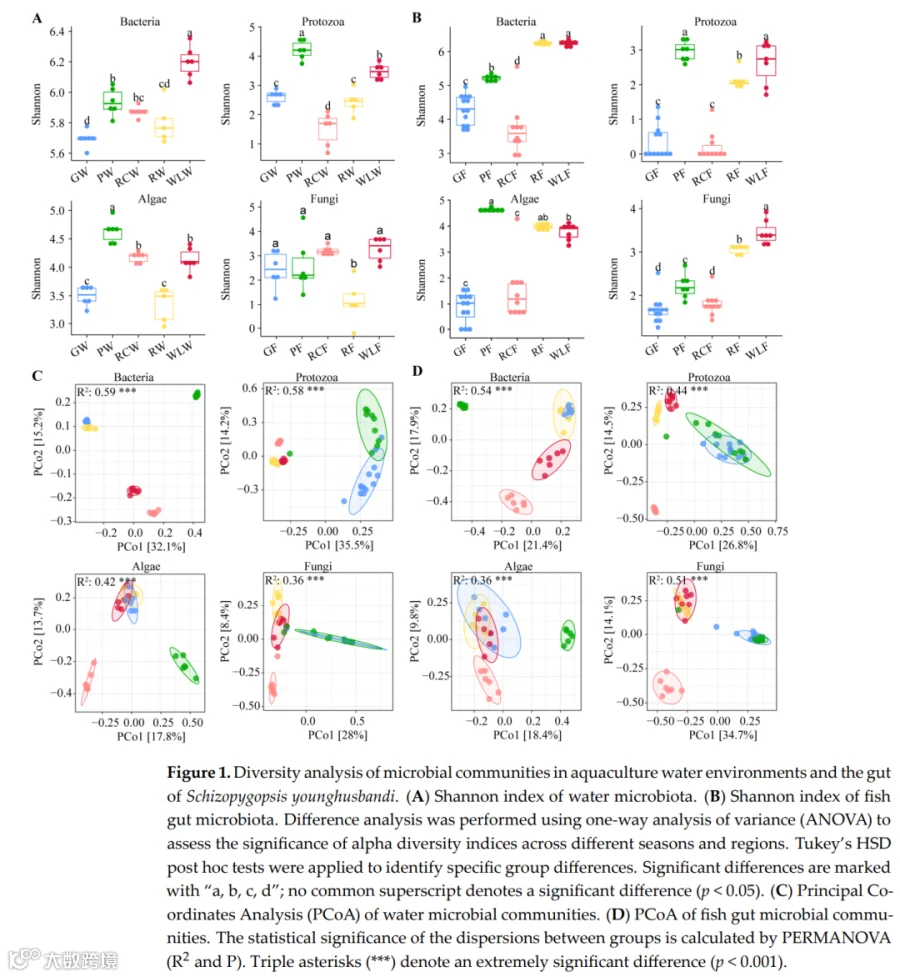
如图1所示,养殖水体和鱼肠道中的微生物α多样性在处理间均存在显著差异。在养殖水体中(图1A),细菌、原生动物、藻类和真菌的Shannon指数均受养殖模式显著影响。细菌多样性在WLW组达到峰值,而原生动物、藻类和真菌多样性在PW组持续最高(p<0.05),表明不同微生物域之间存在强烈的生境特异性分化。在鱼肠道中(图1B),微生物多样性模式在处理间也存在显著差异。细菌多样性在WLF和RF组最高,原生动物多样性在RCF组最低,藻类和真菌多样性在WLF组达到峰值,表明养殖处理导致跨多个微生物类群的肠道微生物多样性发生处理依赖性重组。基于Bray-Curtis距离的PCoA进一步揭示了处理驱动的群落结构变化(图1C,D)。在养殖水体中,细菌群落在处理间表现出最强的分离,表明对环境变化高度敏感,原生动物群落也显示出明显的处理特异性聚类。藻类和真菌群落表现出相对较弱但仍可辨别的分离模式。在肠道微生物群中,细菌和原生动物群落形成了明确的处理簇,而藻类和真菌群落表现出中度但一致的处理相关分化。总之,这些结果表明,不同的养殖模式显著塑造了养殖水体和鱼肠道中的微生物多样性和群落结构,其中细菌和原生动物对环境和养殖管理驱动的变化表现出最高的响应性。
3.养殖水体和鱼肠道中微生物群落的组成及差异
图2总结了不同处理下微生物群落的分类组成和相对丰度。在门水平上,真菌群落以子囊菌门和担子菌门为主,但其相对比例在处理间变化显著,表明强烈的环境和宿主相关效应对真菌群落组装的影响(图2A)。细菌群落主要由假单胞菌门、拟杆菌门和芽孢杆菌门组成(图2B),假单胞菌是大多数样本中的核心和优势组分,而拟杆菌门和芽孢杆菌门呈现处理依赖性富集模式。其他细菌门通常相对丰度较低,并在特定样本中零星富集。藻类群落以绿藻门和硅藻门为主(图2C),在不同处理间呈现相互优势的模式,表明受营养可利用性和光照条件等环境因素的差异调控。原生动物群落主要由纤毛门和丝足虫门组成(图2D),其相对丰度在处理间差异显著,反映了微食物网的处理特异性重构。韦恩分析显示,所有微生物类群中,水体和鱼肠道群落之间存在明显的组成分化(图2E)。尽管有一部分共有类群,但每种生境主要以大量独特物种为特征,特别是在细菌中,共有类群仅占总物种的10.3%,表明水生环境与肠道环境之间存在强烈的生境过滤和生态位分化。
4.微生物群落组成与环境因子的关联
随机森林分析揭示了区分养殖水体处理的关键微生物指示类群(图3A)。关键细菌类群表现出强烈的环境特异性:Flavobacterium在GW组富集,Sphingomonas在WLW组占优势,Limnohabitans与PW组优先相关。藻类和硅藻类群也显示出处理特异性模式,包括Cymbella在GW中富集、Chlorella在PW中富集、Cryptomonas在RCW中富集,而Craticula和Gyrosigma主要与PW组相关。原生动物指示类群同样显示出明显的分布差异,Paramecium在PW中富集,Amoeba在WLW和RCW中丰度更高,表明强烈的环境过滤效应对群落组装的影响。在鱼肠道群落中,随机森林分析识别出处理特异性指示类群(图3B)。Vibrio主要在GF组中富集,而Flavobacterium和Pseudomonas与PF和RCF组相关。Sphingomonas和Bacillus是WLF组的特征类群,伴随明显的藻类、硅藻、真菌和原生生物组合,显示出处理依赖性分化,共同表明养殖模式通过宿主-环境互作重塑了肠道微生物组成。共现网络分析进一步证明了处理依赖性的微生物互作模式重构(图3C)。细菌类群占据中心枢纽位置,并与藻类、真菌和原生动物形成复杂的协同和拮抗相互作用。GW-GF和RCW-RCF处理表现出更高的网络密度和连通性,表明更强的微生物互作、功能互补性和群落稳定性,而WLW-WLF处理显示出降低的连通性,表明网络稳健性和生态稳定性减弱。Spearman相关分析揭示了主要微生物类群与环境变量之间的显著关联(图3D)。细菌群落与TP、TN和DO呈正相关,藻类与pH和DO呈正相关,真菌与NO₂⁻和CaCO₃呈负相关,原生动物与BOD₅和NH₄⁺呈负相关(p<0.05)。这些结果表明,营养可利用性、氧气条件和有机负荷是跨养殖系统塑造跨域微生物群落结构和互作网络的关键驱动因子。RDA结果进一步表明,第一排序轴解释了54.6%的变异,第二轴解释了32.1%(图4)。这两个轴共同解释了肠道微生物群落结构变异的86.7%。总氮、CaCO₃、CaO和水温被确定为显著的解释变量,是驱动群落结构变化的主要因素。
5.养殖水体和鱼肠道中微生物群落的共现网络结构与拓扑特征
共现网络分析揭示了水体和肠道环境中处理依赖性的微生物互作结构差异(图5)。在水体群落中,GW和WLW处理形成了最复杂和连接最紧密的网络,具有更高的节点数、边数和网络密度,表明微生物互作增强、群落整合更强。相比之下,PW和RCW处理表现出简化的网络结构、连接性降低,而RW处理呈现中间互作模式。在肠道群落中,RCF和RF处理表现出最高的网络复杂度,连接性和互作密度显著增加,反映微生物共现和群落整合增强。相比之下,GF和PF处理形成简化网络、互作强度降低,而WLF处理呈现中等网络复杂度。总体而言,肠道微生物网络的结构复杂度低于水体网络,这与肠道环境中的宿主过滤效应和生态位限制一致。拓扑参数分析进一步证实了这些模式。在水体网络中,GW组的节点数和网络密度显著高于所有其他处理(p<0.05),表明其微生物结构连接最紧密,其次是WLW组。PW、RCW和RW组表现出显著较低的网络复杂度。在肠道网络中,RCF和RF组的节点数、边数和网络密度显著高于其他处理(p<0.05),而GF和PF组数值最低,WLF组处于中等水平。正相关和负相关的比例在处理间大致相当,仅有微小变化,表明网络重构主要由互作强度和连通性的变化驱动,而非互作方向的改变。
更多结果和补充图表:doi:10.3390/microorganisms14040925



扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的颠覆性科研数据(0代码生信+统计学)分析平台,同时含有机制图模块(原创3000多素材和机制图模板)+AI一键生成高质量比国自然标书初稿+汉化版Pubmed融合Deepseek高效筛选目标文献同时一键提炼全文核心创新点+SCI文献例句/语料检索模块+全文翻译+文献求助+图片查重+期刊查询+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动发掘更多科研秘籍!