今天给大家解读一篇4月发表在《Stem Cell Research & Therapy》上的题目为“ATG5 overexpression enhances the therapeutic efficacy of mesenchymal stem cells in a mouse colitis model by augmenting anti-inflammatory and antioxidative mechanisms.”的文章。该研究旨在考察ATG5在MSCs抗氧化能力中的作用,并评估ATG5工程化MSCs对结肠炎的治疗效果。通过体外实验(缺氧/血清剥夺预处理、腺病毒介导ATG5过表达)验证ATG5对自噬、抗氧化和增殖的影响,然后在葡聚糖硫酸钠(DSS)诱导的结肠炎小鼠模型中评估疗效,并结合RNA转录组测序、非靶向代谢组学和16S rRNA微生物组分析探索机制。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!包括数据集、绘图代码、图表复现、思路总结、参考文献……0代码!鼠标点点点即可轻松完成5-10分生信SCI全文复现!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!









团队成员合影(位于上海陆家嘴中心,可随时预约参观)





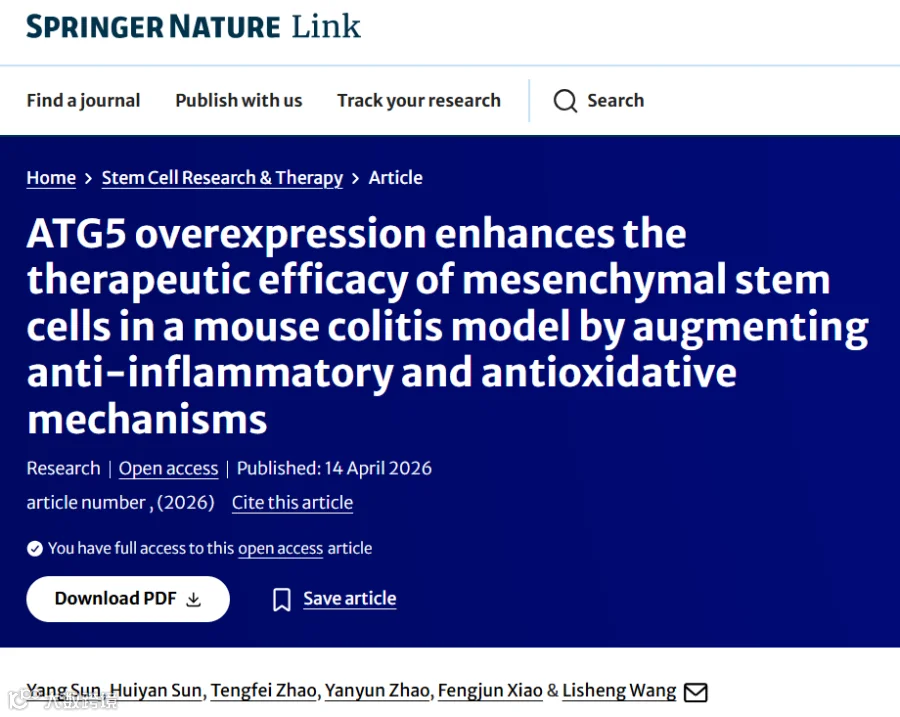
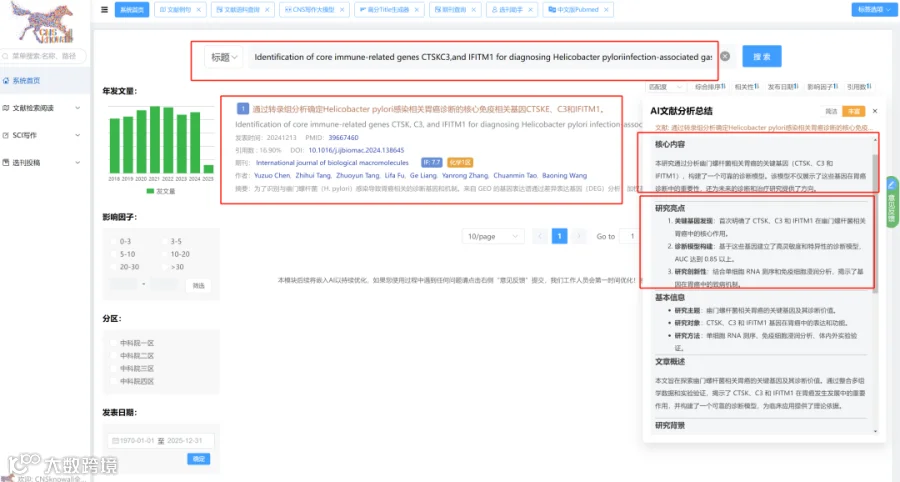
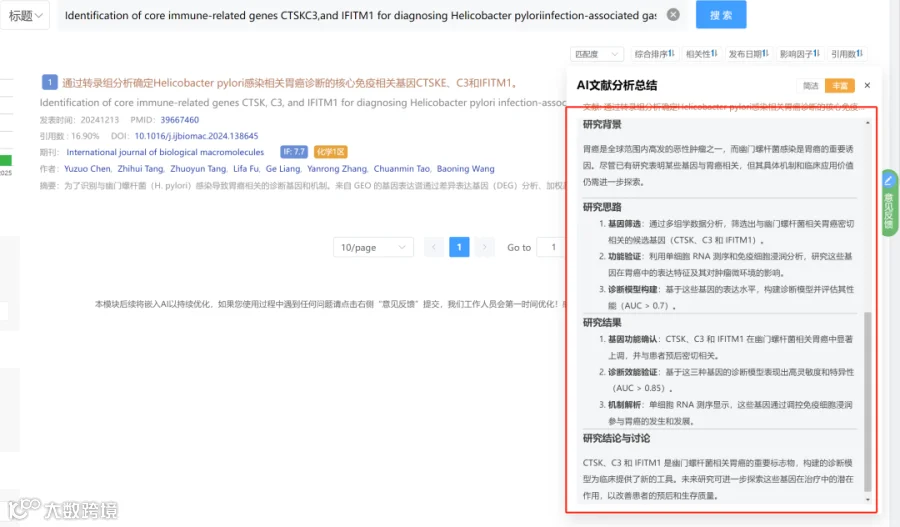
题目:《ATG5过表达通过增强抗炎和抗氧化机制,在小鼠结肠炎模型中提高间充质干细胞的治疗效果》ATG5 overexpression enhances the therapeutic efficacy of mesenchymal stem cells in a mouse colitis model by augmenting anti-inflammatory and antioxidative mechanisms
发表期刊:Stem Cell Research & Therapy
影响因子:7.3
研究背景:
间充质干细胞(MSCs)的治疗效果可通过增强其对炎症微环境的适应性来改善。自噬维持MSCs功能,而自噬相关基因5(ATG5)介导自噬并调节MSCs的生物学功能和治疗效力。此前缺乏关于ATG5在MSCs抗氧化能力中具体作用及其对结肠炎治疗影响的系统研究。
研究思路:
- 体外预实验
用缺氧(1% O₂)和血清剥夺处理MSCs,观察自噬和ATG5表达变化。 - 构建ATG5过表达MSCs(MSCs-ATG5)
通过腺病毒载体实现ATG5过表达,检测自噬活性、抗氧化基因/酶活性、细胞增殖及表面标志物变化。 - 免疫调节功能检测
评估MSCs-ATG5对巨噬细胞极化(M2型)的影响及氧化应激相关信号通路。 - 体内疗效验证
在DSS诱导的结肠炎小鼠中注射MSCs-ATG5,观察疾病症状改善。 - 机制探索
利用转录组分析鉴定被抑制的炎症通路(IL-17/NF-κB),通过代谢组检测结肠组织PGD2水平,通过16S rRNA分析肠道菌群变化。
研究亮点:
-
发现缺氧和血清剥夺预处理可显著增强MSCs的自噬并上调ATG5表达。 -
证实ATG5过表达直接增强MSCs的抗氧化酶活性(谷胱甘肽过氧化物酶、过氧化氢酶)和关键抗氧化基因(HMOX-1、SOD2、CAT)的表达。 -
通过多组学(转录组、代谢组、16S微生物组)揭示ATG5工程化MSCs在结肠炎小鼠中同时调控炎症信号、代谢产物和肠道菌群。
研究结果:
-
缺氧及血清剥夺预处理显著增强MSCs自噬并上调ATG5表达。 -
MSCs-ATG5增强自噬活性和抗氧化能力(上调HMOX-1、SOD2、CAT;增加谷胱甘肽过氧化物酶和过氧化氢酶活性),促进细胞增殖,不改变表面标志物。 -
MSCs-ATG5显著促进M2巨噬细胞极化,调节氧化应激相关信号通路。 -
在小鼠结肠炎模型中,MSCs-ATG5治疗明显改善疾病症状。 -
转录组分析显示MSCs-ATG5抑制IL-17/NF-κB炎症信号通路。 -
代谢组分析表明治疗提高了结肠组织中抗炎代谢物PGD2的水平。 -
微生物组分析发现MSCs-ATG5增加产丁酸菌(如Oscillospirales)的丰度,缓解肠道菌群失调。
研究总结:
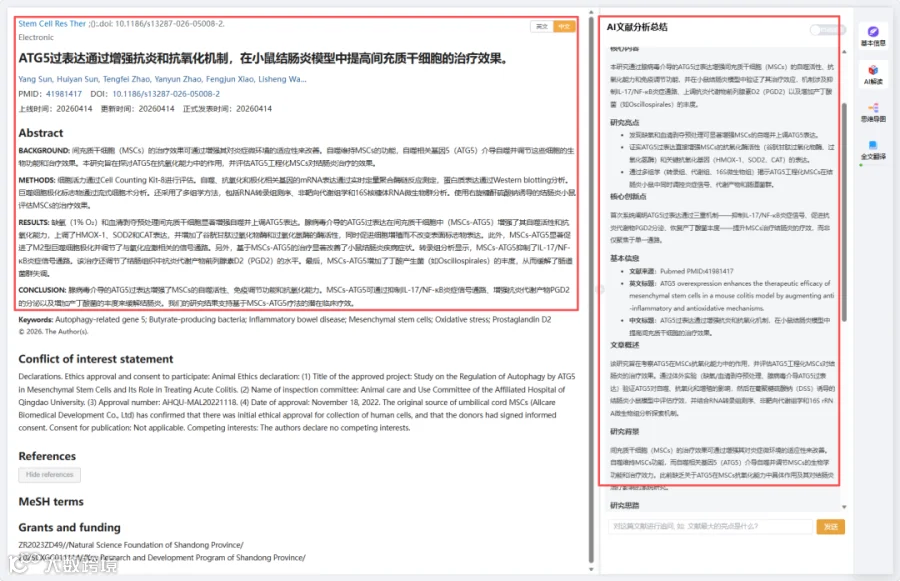
结果译文:
1.缺氧和血清剥夺诱导UC-MSC的自噬
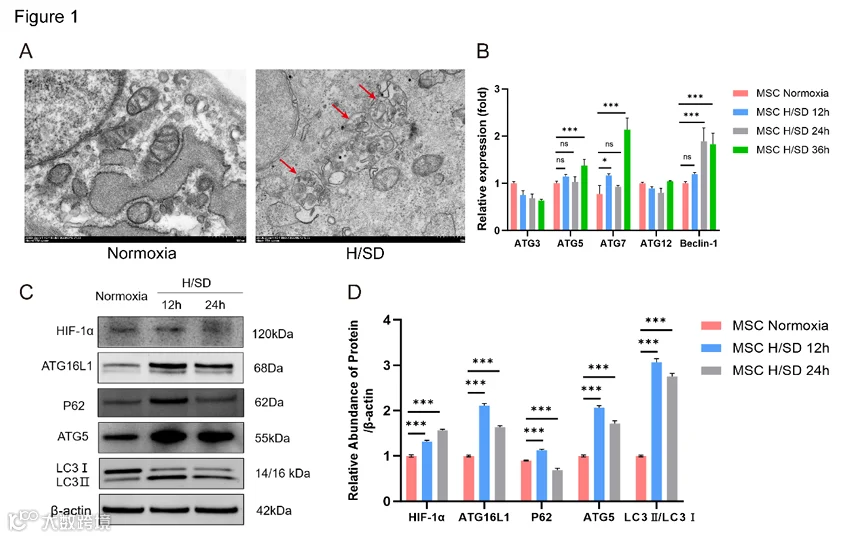
UC-MSC在缺氧(1%氧气)和血清剥夺条件下培养。图1A显示了H/SD条件下24小时自噬体的形成。随后,我们在MSC于H/SD条件下孵育12、24和36小时后,通过qRT-PCR检测了经典自噬相关基因(包括ATG3、ATG5、ATG7、ATG12和Beclin-1)的表达水平。与常氧条件相比,H/SD条件显著增强了ATG5、ATG7和Beclin-1的表达(图1B)。此外,基于蛋白质印迹分析,H/SD条件下12小时和24小时HIF-1α、ATG16L1、ATG5和LC3 II/I的蛋白表达均上调。P62表达在12小时上调,随后在24小时下调,表明H/SD条件下自噬流畅通无阻(图1C-D)。
2.转染Ad-ATG5的MSC的生物学特性
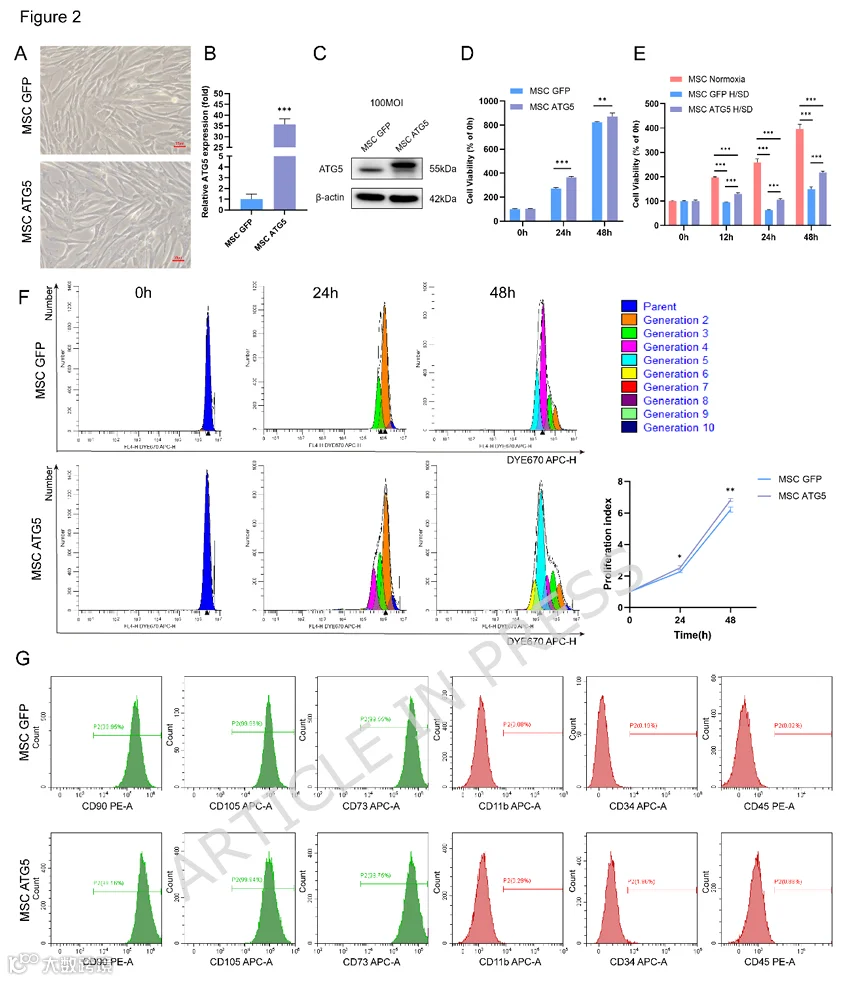
携带ATG5的腺病毒载体示意图见图S1A。将Ad-GFP和Ad-ATG5腺病毒颗粒分别以MOI 100和200转染UC-MSC。转染效率在MOI 100时达到78.7%,在MOI 200时达到92.8%(图S1B、C)。MSC转染Ad-GFP或Ad-ATG5后通过荧光显微镜观察到GFP荧光(图2B)。qRT-PCR和蛋白质印迹分析证实,MSCs-ATG5中ATG5在mRNA和蛋白水平均过表达(图2C、D)。通过流式细胞术检测了MSC-GFP和MSC-ATG5表面标志物(如CD90、CD105、CD73、CD11b、CD34和CD45)的表达。腺病毒介导的ATG5转染未改变细胞形态或MSC表面标志物的表达(图2A、G)。然而,在常氧和H/SD条件下,转染后24和48小时,MSCs-ATG5的细胞活力均显著高于MSCs-GFP(图2D、E)。类似地,Dye670染色分析表明,与MSCs-GFP相比,MSCs-ATG5表现出显著更高的增殖指数(图2F)。
3.MSCs-ATG5促进巨噬细胞的M2极化
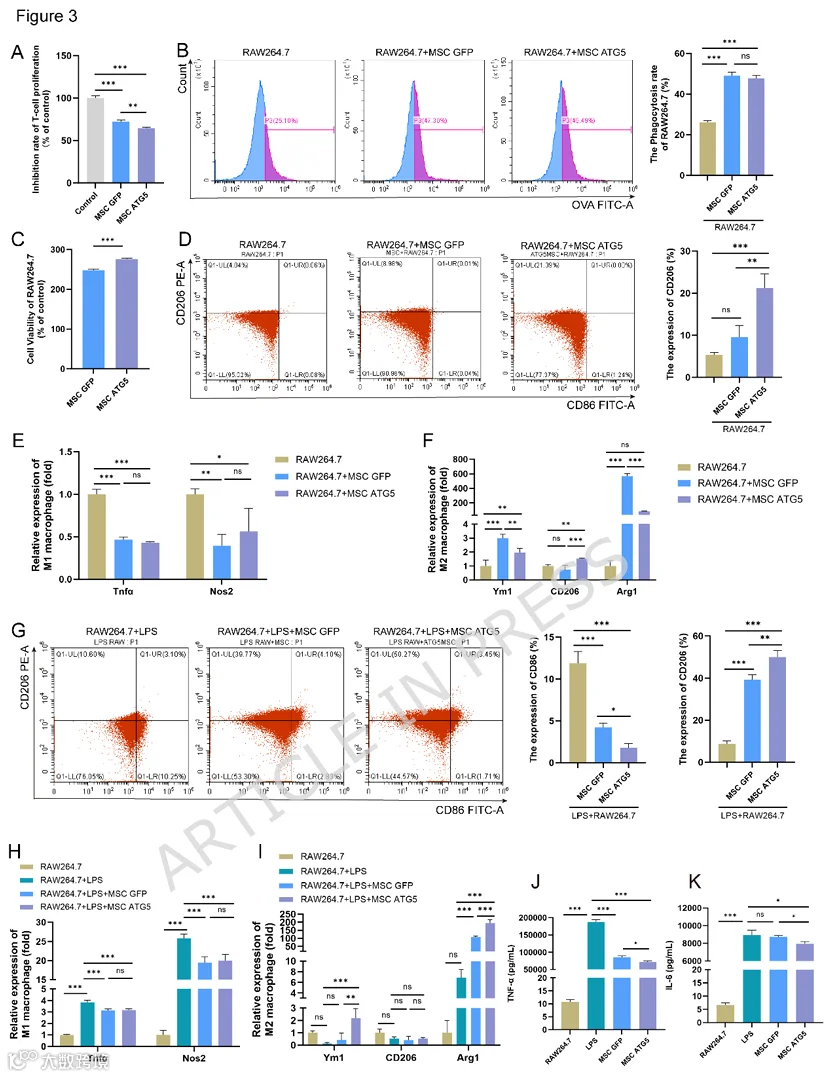
在本部分中,T细胞和巨噬细胞被用作效应细胞来验证MSC的免疫调节能力。荧光素酶-T实验表明,与MSCs-GFP相比,MSCs-ATG5在抑制T细胞增殖方面具有显著增强的功效(图3A)。RAW264.7细胞通常用作巨噬细胞极化的模型。因此,将MSCs-GFP和MSCs-ATG5分别与RAW264.7细胞共培养24小时。通过流式细胞术检测吞噬能力。共培养24小时后,MSCs-GFP和MSCs-ATG5均显著增强了RAW264.7细胞的吞噬能力,但在MSCs-ATG5和MSCs-GFP之间未观察到吞噬增强效应的显著差异(图3B)。MSCs-ATG5对RAW264.7细胞增殖的增强作用强于MSCs-GFP(图3C)。使用流式细胞术检测M1巨噬细胞标志物CD86和M2巨噬细胞标志物CD206。将RAW264.7细胞与MSC共培养后,RAW264.7细胞的M2极化能力显著增强,且MSCs-ATG5对RAW264.7细胞M2极化(CD206+)的增强作用更强(图3D)。与MSC共培养后,通过qRT-PCR检测M1/M2巨噬细胞标志物。MSCs-ATG5抑制了部分M1标志物(如Tnfa和Nos2)的表达,并促进了M2标志物(如Ym1、CD206和Arg1)的表达(图3E、F)。此外,使用LPS诱导巨噬细胞炎症。LPS显著增加了M1巨噬细胞(CD86+)的比例(图3G)。将LPS诱导的RAW264.7细胞与MSC共培养后,MSCs-ATG5具有更强的抑制M1巨噬细胞的能力,并能促进巨噬细胞向M2巨噬细胞转化(图3G)。qRT-PCR结果显示,LPS显著促进了M1巨噬细胞标志物(如Tnfa和Nos2)的表达,并抑制了M2巨噬细胞标志物(如Ym1、CD206和Arg1)的表达(图3H、I)。qRT-PCR结果与MSCs-ATG5共培养后的流式细胞术结果一致。两种MSC(MSCs-GFP和MSCs-ATG5)均抑制了M1巨噬细胞标志物(Tnfa和Nos2)的表达,同时上调了M2巨噬细胞标志物(Ym1和Arg-1)的表达。与MSCs-GFP组相比,MSCs-ATG5组在M1标志物表达方面无显著差异,但显著增强了M2标志物的表达。LPS显著增强了RAW264.7细胞分泌促炎细胞因子,包括TNF-α和IL-6。与MSCs-GFP相比,MSCs-ATG5对这些细胞因子分泌的抑制能力增强(图3J、K)。
4.ATG5调节MSC中的氧化应激信号通路
为了进一步阐明ATG5调节MSC生物学特性的机制,对MSCs-GFP和MSCs-ATG5进行了RNA测序,每组三个生物学重复。基于两组之间鉴定的DEGs,进行了KEGG富集分析以探索MSCs-GFP和MSCs-ATG5之间的信号通路和功能差异。根据与DEGs相关的显著改变的代谢和信号转导通路(MSCs-ATG5 vs MSCs-GFP),p53信号通路、细胞衰老和铁死亡主要富集(图4B)。值得注意的是,这些信号通路与氧化应激相关。通过筛选DEGs并将其映射到这些特定信号通路,关键DEGs可总结如下。在p53信号通路中,MSC中ATG5过表达显著增加了FAS、SESN2、SESN3、CDKN1A、IGFBP3和MDM2的表达。在细胞衰老信号通路中,MSC中ATG5过表达显著增加了PIK3R3、CDKN1A、IGFBP3和MDM2的表达。在铁死亡信号通路中,MSC中ATG5过表达显著增加了SLC7A11的表达。
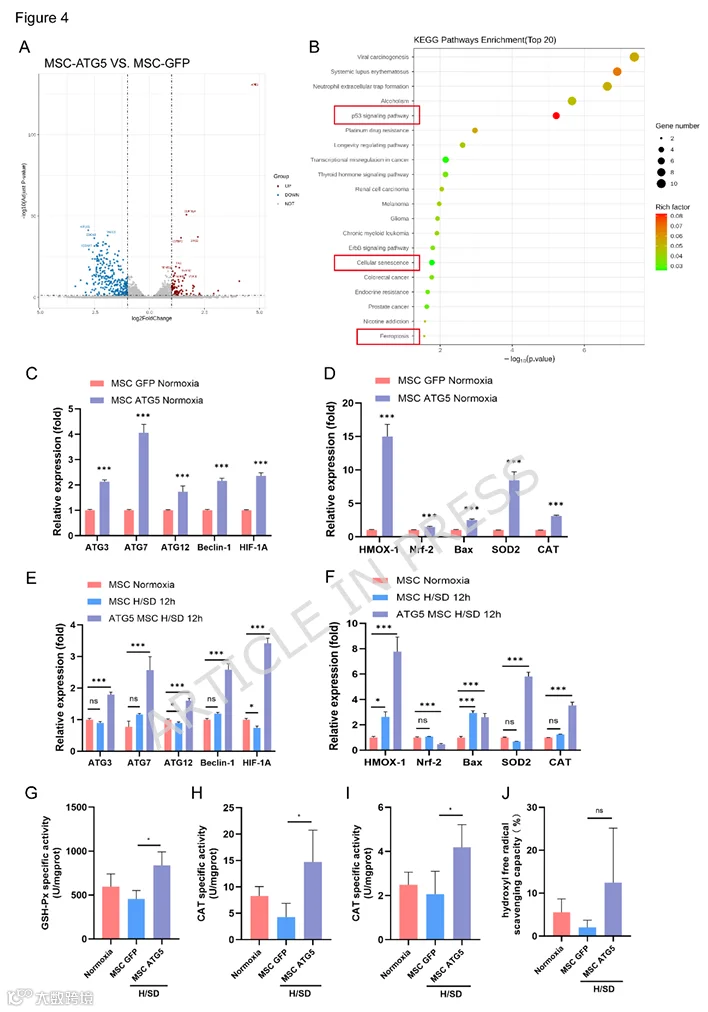
5.ATG5过表达上调MSC中自噬和抗氧化应激相关基因的表达
6.MSCs-ATG5治疗减轻DSS诱导的小鼠结肠炎
为了评估MSCs-ATG5的体内治疗效果,我们使用了DSS诱导的急性溃疡性结肠炎小鼠模型。实验流程如图5A所示。DSS诱导后,小鼠体重在第5天开始下降,而MSCs-ATG5治疗在DSS+PBS组和DSS+MSCs-GFP组体重持续下降的情况下,显著恢复了体重(图5B)。与NC组相比,DSS+PBS组结肠长度显著缩短;MSCs-ATG5治疗有效恢复了结肠长度,而MSCs-GFP治疗未产生显著效果(图5C、D)。疾病活动指数(DAI)分析显示,与DSS+PBS组相比,MSCs-ATG5治疗显著降低了DAI评分(图5E)。结肠组织的H&E染色显示,与DSS+PBS组相比,MSCs-ATG5治疗显著减轻了炎症细胞浸润、隐窝损伤和黏膜下层水肿(图5F)。Luminex检测显示,与DSS+PBS组相比,MSCs-ATG5治疗降低了促炎细胞因子(包括IL-1β、TNF-α、IFN-γ、IL-6、G-CSF和IL-17A)的血清水平,同时增加了抗炎因子如IL-10和IL-4的分泌(图5G)。此外,MSC-ATG5治疗上调了肠上皮紧密连接蛋白occludin和ZO-1的表达(图5H、J、K)。值得注意的是,免疫荧光显示MSCs-ATG5治疗组结肠组织中CD206+巨噬细胞上调(图5I)。
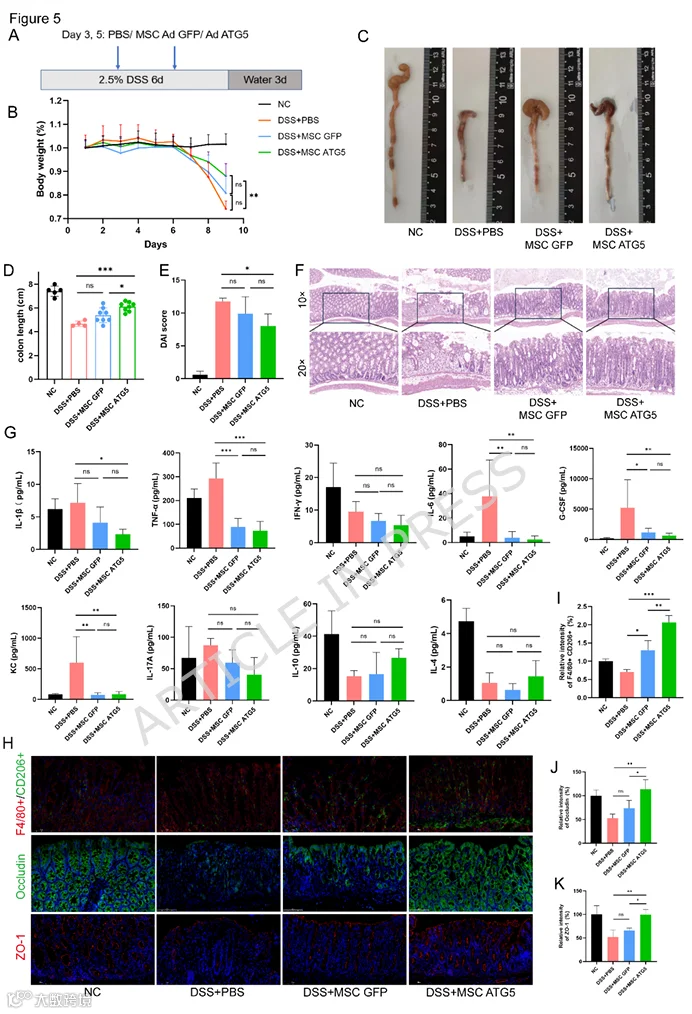
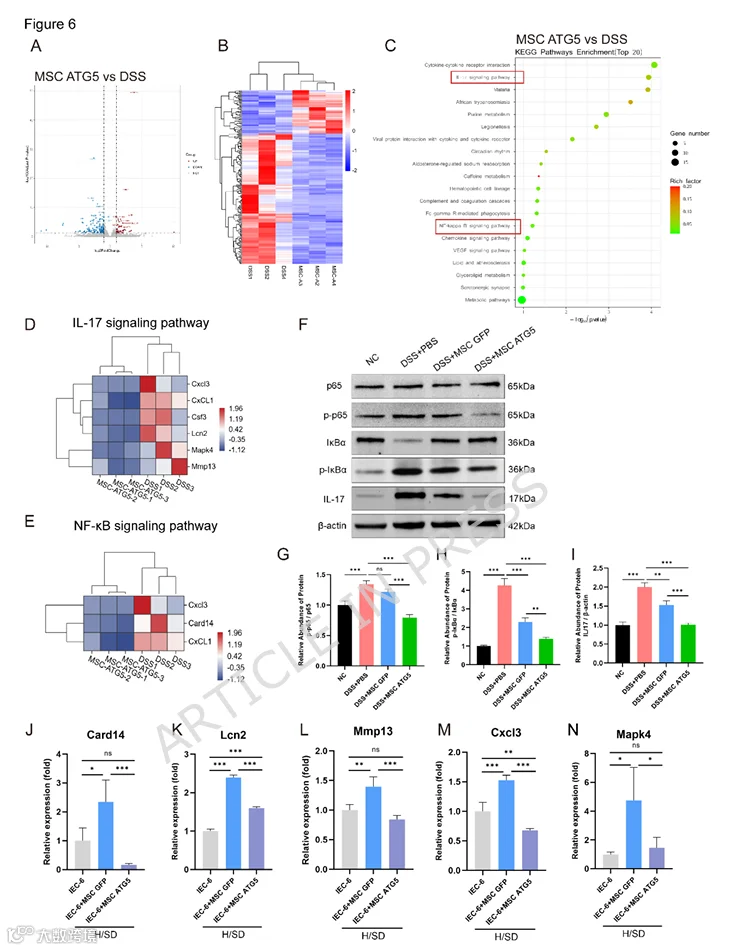
7.MSC-ATG5治疗抑制DSS诱导结肠炎小鼠的IL-17/NF-κB炎症信号通路
对上述小鼠模型的结肠组织进行了RNA-seq分析。随后的KEGG富集分析显示,与正常组相比,DSS模型组中上调的基因主要富集在经典的炎症相关信号通路中。这些通路包括IL-17信号通路、TNF信号通路以及与IBD相关的通路(图S2D)。随后,我们分析了MSC-ATG5组和DSS模型组之间的DEGs。图6A、B显示了ATG5组与DSS组相比DEGs的火山图和热图。有趣的是,KEGG富集分析显示,MSC-ATG5组中下调的基因在IL-17和NF-κB信号通路中显著富集(图6C)。其中,被MSCs-ATG5抑制的IL-17信号通路中的基因包括Csf2、Mapk4、Cxcl3、Cxcl1、Mmp13和Lcn2(图6D),NF-κB信号通路中下调的基因包括Card14、Cxcl3和Cxcl1(图6E)。在DSS诱导的小鼠结肠炎中,NF-κB信号通路被显著激活,表现为p65蛋白磷酸化增加、IkBα蛋白降解以及IL-17信号通路蛋白上调。MSC-ATG5治疗有效降低了p65磷酸化,增强了IkBα蛋白稳定性,并抑制了IL-17信号通路(图6F-I)。接下来,我们在体外进行了IEC-6和MSC的共培养实验。使用H/SD条件模拟体内的氧化应激环境。通过qRT-PCR检测上述IL-17和NF-κB信号通路基因。结果表明,MSCs-ATG5能够抑制Card14、Mmp13和Cxcl3等基因的表达(图6J-N)。对DEGs的GO富集分析显示,MSCs-ATG5调节结肠细胞中的多个生物学过程,如免疫反应、免疫系统过程、B细胞介导的免疫和炎症反应(图S2E)。
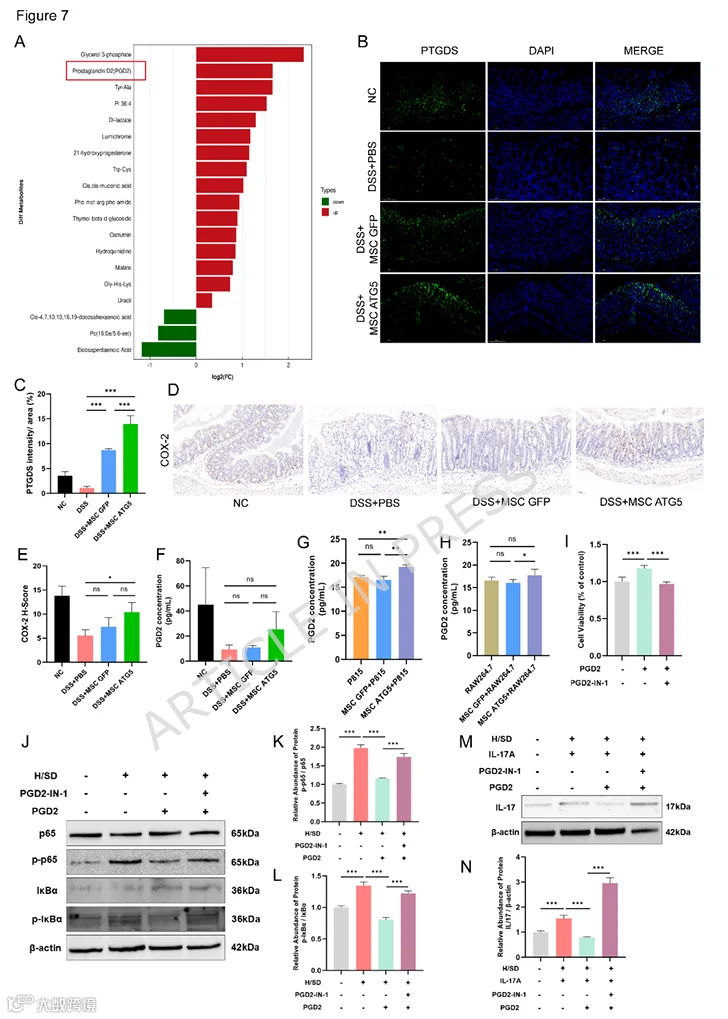
8.MSCs-ATG5调节结肠组织中的抗炎代谢物PGD2
溃疡性结肠炎中的炎症相关代谢物是调节其病理进展的关键因素。我们对MSC-ATG5组和DSS模型组小鼠的结肠组织进行了非靶向代谢组学测序。火山图显示了通过正/负离子模式筛选鉴定的差异代谢物(MSCs-ATG5 vs. DSS,OPLS-DA VIP > 1,P < 0.05)。基于正/负离子模式的具体差异代谢物以蝴蝶图说明。蝴蝶图显示了ATG5组中上调和下调的代谢物,以及差异比率的分析。我们选择负离子模式下的PGD2用于后续实验(图7A)。环氧合酶-2(COX-2)和PGD2通过体内的前列腺素合成途径具有密切的上下游关系。因此,我们使用免疫荧光染色检测了结肠组织中PGD2合酶(PTGDS)的表达(图7B、C),结果表明MSC-ATG5治疗显著增加了PTGDS水平。随后,我们检测了结肠组织中COX-2的表达,结果显示与其他组相比,MSC-ATG5组COX-2表达呈上调趋势(图7D、E)。为了进一步研究MSCs-ATG5如何调节前列腺素PGD2,我们量化了小鼠血清中该蛋白的水平,结果显示MSCs-ATG5组中PGD2水平升高(图7F)。此外,我们在常氧和H/SD条件下测量了MSC和MSCs-ATG5分泌PGD2的情况。MSC在基础条件下分泌低水平的PGD2,而ATG5过表达显著增加了PGD2分泌(图S3D)。值得注意的是,随着H/SD条件暴露时间的延长,PGD2分泌能力下降(图S3E)。在H/SD条件下孵育后,MSCs-ATG5的培养上清液(图S3F)和细胞内(图S3G)中的PGD2水平均显著高于MSCs-GFP。PGD2主要由肥大细胞和巨噬细胞分泌。因此,我们将肥大细胞系P815和巨噬细胞系RAW264.7与MSCs-GFP或MSCs-ATG5共培养。MSCs-ATG5显著促进了P815和RAW264.7细胞分泌PGD2(图7G、H)。更重要的是,PGD2促进了IEC-6细胞的增殖(图7I)。在H/SD条件下,PGD2对IEC-6细胞中的NF-κB和IL-17信号通路具有抑制作用,而这种抑制作用被PGD2拮抗剂(PGD2-IN-1)逆转(图7J-N)。
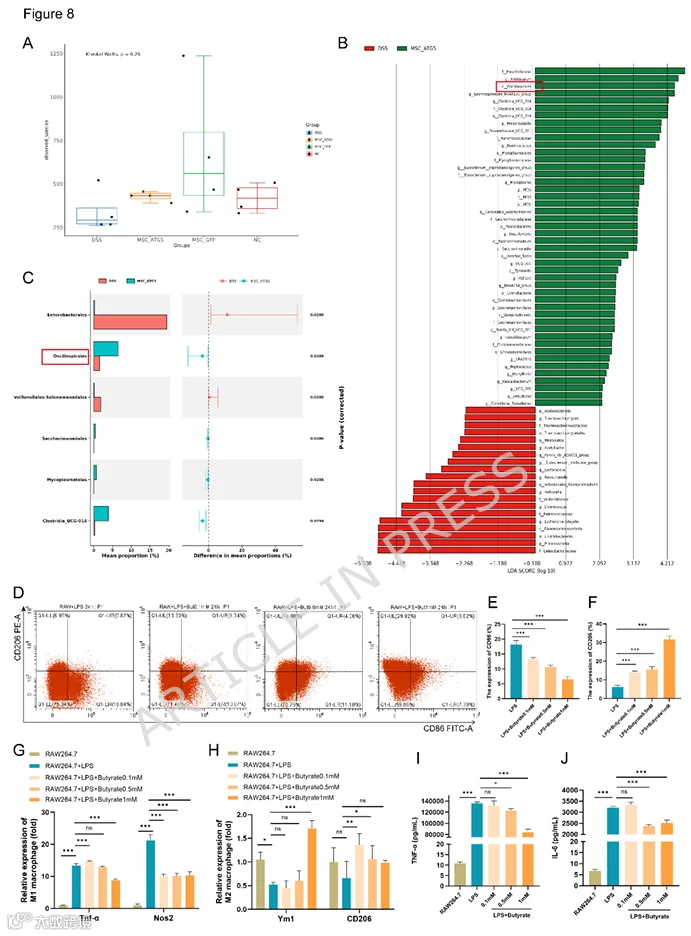
9.MSCs-ATG5改变肠道菌群组成并增加产丁酸菌的比例
MSC-ATG5治疗后,DSS诱导的肠道菌群组成发生显著改变(图8A)。使用LEfSe方法鉴定MSC-ATG5组和DSS组之间存在显著差异的微生物生物标志物。通过LEfSe数据的可视化,该图显示了MSC-ATG5组和DSS组之间显著差异物种的LDA评分。MSC-ATG5治疗后,肠道菌群中益生菌类群如普雷沃氏菌科、Allobaculum和Oscillospirales的相对丰度显著高于DSS-PBS治疗后(图8B)。Oscillospirales包含产丁酸细菌。使用STAMP差异分析方法比较MSC-ATG5组和DSS组之间的物种丰度。类似地,MSC-ATG5组中Oscillospirales的平均比例更高(图8C)。通过对肠道菌群的KEGG功能分析,MSC-ATG5治疗显著增强了肠道菌群对环境变化的代谢功能适应性,包括复制和修复、细胞生长和耐药性(图S4C)。流式细胞术结果显示,与DSS组相比,MSC-ATG5组中CD206+巨噬细胞的比例更高。丁酸钠(0.1、0.5和1 mM)以浓度依赖性方式增加了CD206+巨噬细胞的比例,并降低了CD86+巨噬细胞的比例(图8D-F)。qRT-PCR分析显示,丁酸钠抑制了M1标志物(Tnfa、Nos2)的表达,并促进了M2标志物(Ym1、CD206)的表达(图8G)。此外,丁酸钠还抑制了LPS刺激的RAW264.7细胞分泌TNF-α和IL-6(图8I、J)。这些结果表明,丁酸促进巨噬细胞向M2表型极化,并抑制炎症反应。
更多结果和补充图表:doi:10.1186/s13287-026-05008-2



扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的颠覆性科研数据(0代码生信+统计学)分析平台,同时含有机制图模块(原创3000多素材和机制图模板)+AI一键生成高质量比国自然标书初稿+汉化版Pubmed融合Deepseek高效筛选目标文献同时一键提炼全文核心创新点+SCI文献例句/语料检索模块+全文翻译+文献求助+图片查重+期刊查询+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动发掘更多科研秘籍!