胰岛移植是治疗1型糖尿病的有效手段,但移植后胰岛早期凋亡严重影响疗效。本研究探索了凋亡骨髓间充质干细胞(BMSCs)对移植胰岛的保护作用。通过星形孢菌素(STS)诱导BMSCs凋亡(凋亡率约41%),将其与胰岛共移植于糖尿病大鼠肾被膜下,发现BMSCs-STS组血糖在24小时内恢复正常,比对照组更快,且移植物存活率更高、凋亡减少。体外实验中,凋亡BMSCs条件培养基(CM)可显著抑制STZ诱导的INS-1细胞凋亡。蛋白质组学分析鉴定出201个差异表达蛋白,其中铁负载型Lcn2(holo-Lcn2)是关键抗凋亡因子。机制研究表明,holo-Lcn2通过结合细胞膜受体Slc22a17,将Fe³⁺转运入细胞内,提高Bcl-2/Bax比率,抑制细胞凋亡。敲低Slc22a17则消除保护作用。本研究揭示了凋亡BMSCs通过holo-Lcn2/Slc22a17/Fe³⁺轴减少移植胰岛凋亡的新机制,为胰岛移植的无细胞疗法提供了新靶点。
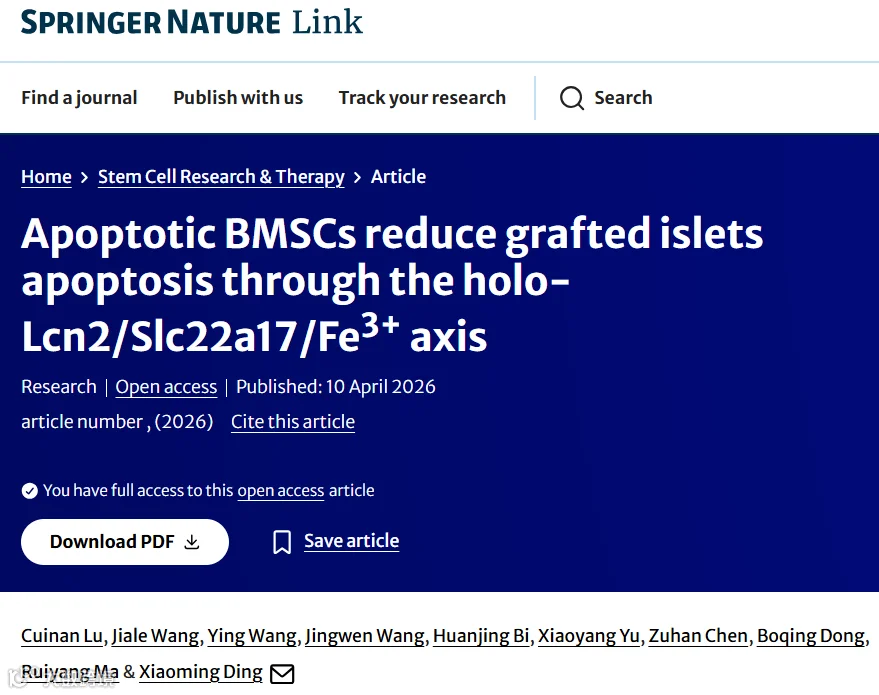
今天给大家解读一篇4月发表在《Stem Cell Research & Therapy》上的题目为“Apoptotic BMSCs reduce grafted islets apoptosis through the holo-Lcn2/Slc22a17/Fe3+ axis.”的文章。该研究针对移植胰岛早期凋亡这一关键挑战,采用Staurosporine预诱导BMSCs凋亡,收集条件培养基预处理β细胞,显著抑制了β细胞凋亡。体内实验将胰岛与凋亡BMSCs共移植至糖尿病大鼠肾包膜下,有效抑制胰岛凋亡并改善移植结局。蛋白质组学分析发现,凋亡BMSCs分泌的holo-Lcn2是关键效应分子,它结合Slc22a17转运体促进Fe³⁺内流;抑制Fe³⁺转运则削弱其抗凋亡作用。(请持续关注我们,每天为您解读最新见刊的文献!)想薅生信资料羊毛?直接在对话框回复 “资料”,免费领取干货大礼包!包括数据集、绘图代码、图表复现、思路总结、参考文献……0代码!鼠标点点点即可轻松完成5-10分生信SCI全文复现!
不想做实验,没数据,还想要快速发表文章,没问题的!公共数据库就是我们的数据宝藏!没思路不用担心,作为专业的生信团队,我们很乐意为你们效劳,提供研究路线设计和数据挖掘分析,扫码联系我们吧!



团队成员合影(位于上海陆家嘴中心,可随时预约参观)
题目:《凋亡的BMSCs通过holo-Lcn2/Slc22a17/Fe3+轴减少移植胰岛的凋亡》Apoptotic BMSCs reduce grafted islets apoptosis through the holo-Lcn2/Slc22a17/Fe3+ axis
发表期刊:Stem Cell Research & Therapy
影响因子:7.3
研究背景:
移植胰岛的早期凋亡是严重影响胰岛移植疗效的关键挑战之一,亟待探寻新的保护策略。BMSCs已被广泛研究,但其凋亡状态下的旁分泌作用尚不明确。
CNSknowall 平台 Pubmed+AI 快速提炼全文要点
研究思路:
-
体外:用Staurosporine诱导BMSCs凋亡,收集条件培养基处理β细胞,观察凋亡抑制效果。
-
体内:将胰岛与凋亡BMSCs共移植于糖尿病大鼠肾包膜下,评估胰岛存活与移植效果。
-
机制:通过蛋白质组学筛选差异蛋白,鉴定holo-Lcn2,并验证其通过Slc22a17转运Fe³⁺进入细胞发挥抗凋亡作用。
研究亮点:
-
首次发现凋亡诱导的BMSCs条件培养基能显著抑制β细胞凋亡,且共移植实验验证了体内保护效应。
-
通过蛋白质组学锁定关键分子holo-Lcn2,并阐明其通过Slc22a17/Fe³⁺轴发挥抗凋亡作用,为无细胞疗法提供了新机制。
研究结果:
-
凋亡BMSCs的条件培养基显著增强了对β细胞凋亡的抑制。
-
共移植凋亡BMSCs的糖尿病大鼠胰岛凋亡减少,移植结局更佳。
-
蛋白质组学显示凋亡BMSCs分泌的holo-Lcn2具有抗凋亡活性;holo-Lcn2结合Slc22a17促进Fe³⁺内流,抑制Fe³⁺转运则消除其保护效应。
研究总结:
凋亡BMSCs通过holo-Lcn2/Slc22a17/Fe³⁺轴减少移植胰岛的凋亡,该机制为基于BMSCs的无细胞疗法在胰岛移植中的应用提供了新见解。未来可进一步探索该通路在临床转化中的潜力。
结果译文:
采用全骨髓贴壁法成功分离BMSCs后,P3代BMSCs在光学显微镜下呈现典型的梭形形态(图S1B)。通过茜素红染色和油红O染色分别证实了BMSCs向成骨细胞和脂肪细胞的分化潜能(图S1C-D)。流式细胞术表征了BMSCs的表面标志物,结果显示CD90和CD29阳性表达,CD45和CD11b/c阴性表达(图S1E)。
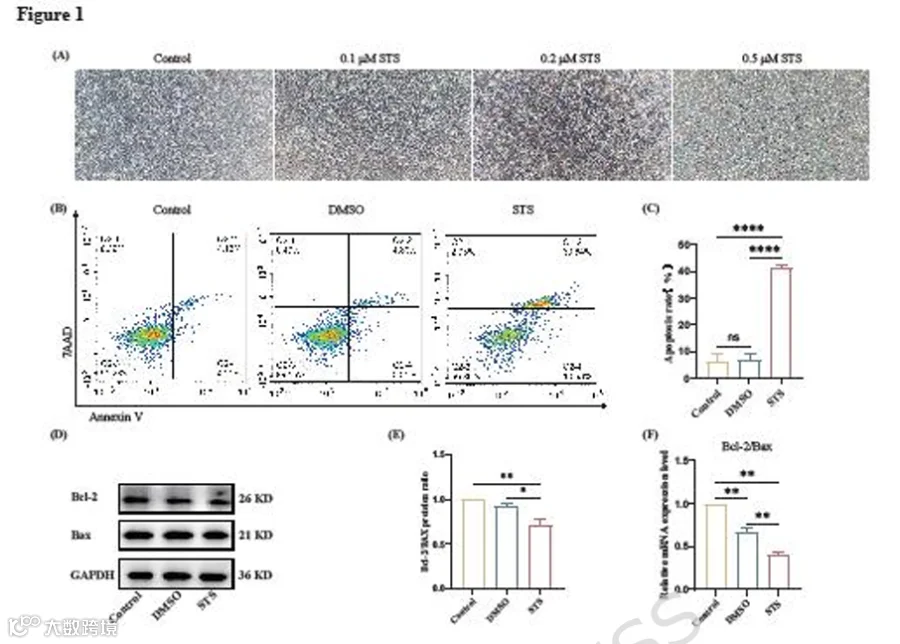
为建立凋亡BMSCs模型,我们用不同浓度的STS处理BMSCs,并在光学显微镜下观察其形态。随着STS浓度增加,BMSCs的形态逐渐变得更加细长。值得注意的是,当浓度达到0.5 μM处理2小时时,细胞出现明显的漂浮和死亡现象(图1A)。因此,我们选择0.2 μM STS处理2小时来诱导BMSCs凋亡。流式细胞术结果显示,STS处理组凋亡细胞(Annexin V阳性)比例为41.47% ± 3.16%,而DMSO对照组为5.13% ± 2.12%(图1B-C)。Western blot结果显示,与对照组相比,STS处理组Bcl-2蛋白表达降低,Bax蛋白表达升高(图1D-E)。RT-qPCR结果与蛋白水平一致(图1F)。这些结果表明,0.2 μM STS处理2小时成功诱导了BMSCs凋亡。
2.BMSCs-STS与胰岛细胞共移植改善了胰岛移植的治疗效果
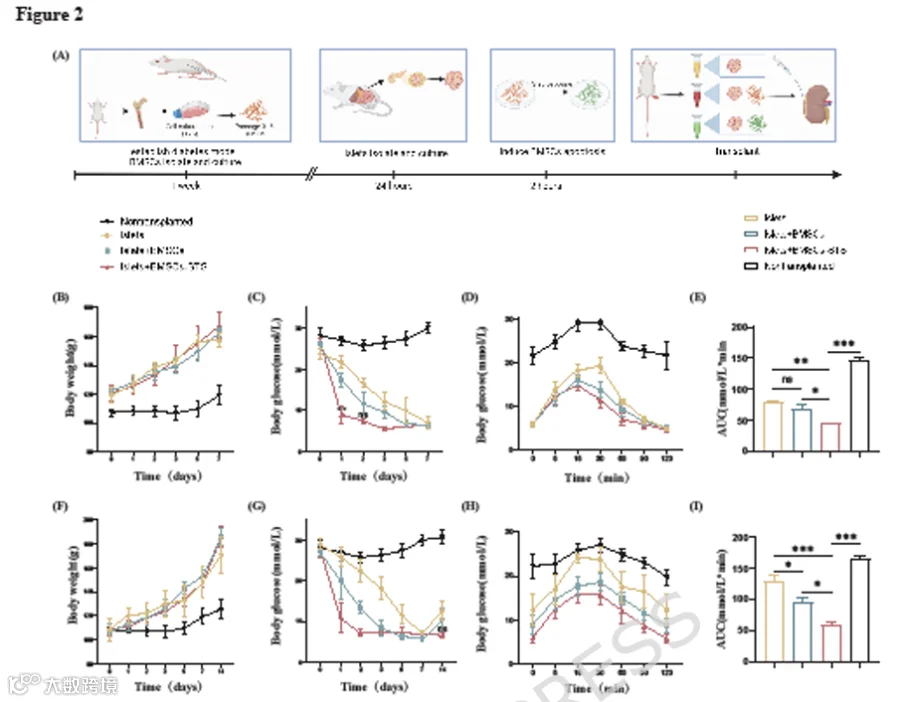
MSCs与胰岛共移植可增强胰岛移植的治疗效果,且部分研究表明凋亡BMSCs在其他疾病模型中可能具有更好的治疗效果。为了评估STS诱导的凋亡BMSCs与胰岛共移植的体内效应,并验证与活BMSCs相比,BMSCs-STS是否能改善移植胰岛的存活,我们设计了如下实验(图2A):STZ诱导的糖尿病大鼠随机分为四组(n=5):未移植组、仅胰岛移植组(Islets)、胰岛与BMSCs共移植组(Islets+BMSCs)以及胰岛与BMSCs-STS共移植组(Islets+BMSCs-STS)。各组细胞被移植到糖尿病大鼠肾被膜下,并在术后第7天和第14天取出移植物进行观察。
我们在术后第1、2、3、5、7和14天检测了大鼠的随机血糖和体重。所有移植组大鼠的体重逐渐增加,无显著差异,而未移植大鼠体重增长缓慢甚至下降(图2B、F)。移植后,仅Islets+BMSCs-STS组大鼠在24小时内血糖即恢复正常,随后保持稳定。此时,与Islets+BMSCs组相比,血糖水平存在显著差异(P<0.05)。Islets+BMSCs组在术后第3天血糖恢复正常,而Islets组约在第5天血糖恢复正常(图2C、G)。然而,Islets组部分大鼠出现血糖反弹。
为了进一步评估移植物功能,我们在术后第7天和第14天对四组大鼠进行了腹腔葡萄糖耐量试验。所有胰岛移植组的血糖曲线下面积均显著降低(图2D-E、H-I),其中Islets+BMSCs-STS组AUC最小,表明其葡萄糖处理能力最佳。
3.BMSCs-STS与胰岛细胞共移植增加了移植物存活并减少移植物凋亡
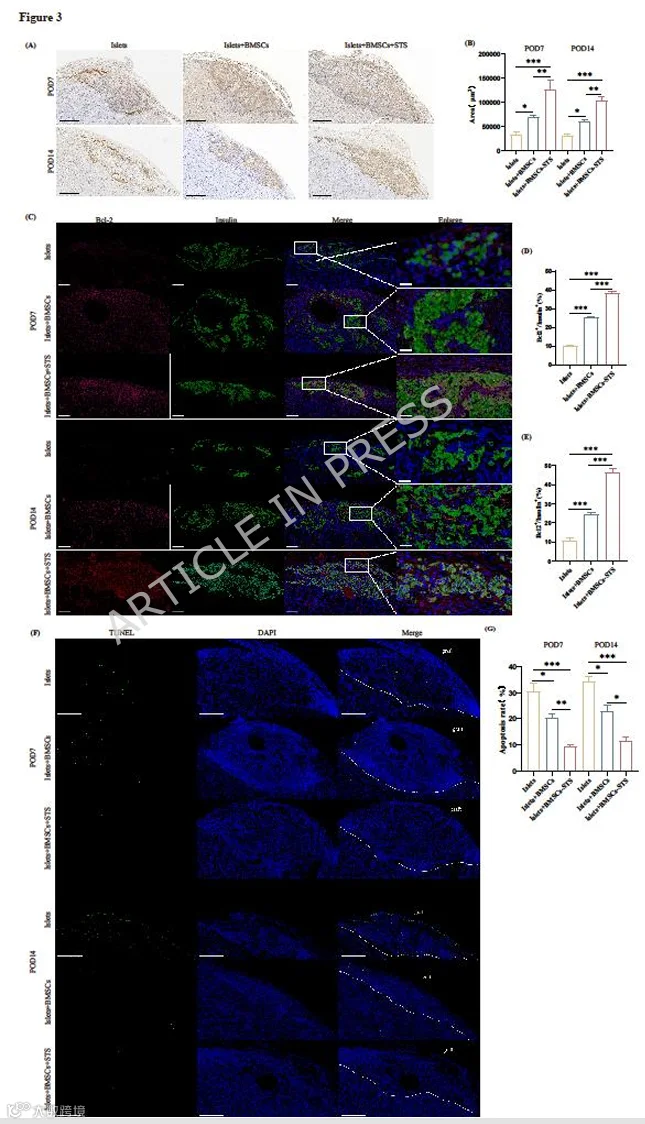
为了进一步明确BMSCs-STS是否能减少早期移植物凋亡并提高移植物存活率,我们在术后第7天和第14天取出移植物,进行了胰岛素免疫组化染色以及Bcl-2和胰岛素双免疫荧光染色。胰岛素免疫组化染色显示,Islets+BMSCs-STS组在所有移植组中具有最大的胰岛素阳性面积,表明更高的移植物存活率和存活的移植物具有更好的胰岛素分泌功能(图3A-B)。Bcl-2和胰岛素双免疫荧光染色可评估移植物凋亡,结果显示与Islets组和Islets+BMSCs组相比,Islets+BMSCs-STS组具有最大的Bcl-2+/胰岛素+染色面积,表明Islets+BMSCs-STS组凋亡显著减少(图3C-E)。通过TUNEL染色进一步评估了移植胰岛的凋亡。成像分析与Bcl-2+/胰岛素+染色结果一致,显示Islets+BMSCs-STS组凋亡水平最低(图3F-G)。这些结果共同表明,BMSCs-STS共移植可减少移植物凋亡并提高胰岛移植物存活率。
4.BMSCs-STS条件培养基保护胰岛和β细胞免于凋亡
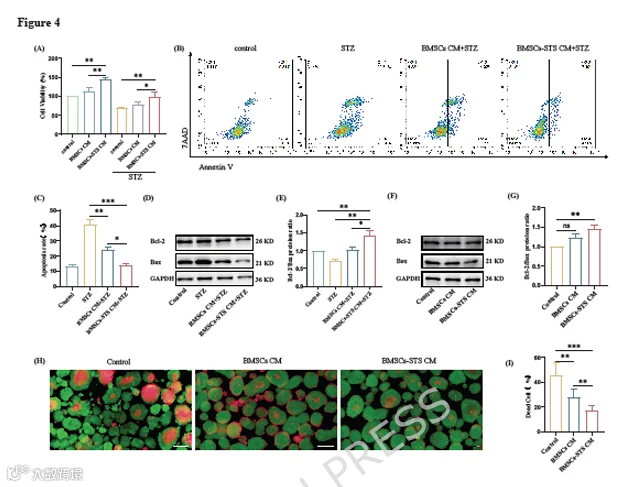
为了在体外评估BMSCs-STS的抗凋亡功能,我们收集了BMSCs-STS CM和BMSCs CM,并用它们预处理INS-1细胞和胰岛。
如前所述,3 mM STZ是处理INS-1细胞的最佳浓度。因此,我们首先用两种CM预处理INS-1细胞24小时,然后用3 mM STZ处理24小时。通过CCK-8实验评估INS-1细胞活力。我们发现BMSCs-STS CM预处理显著增强了INS-1细胞的活力(图4A)。随后,我们使用Western blot、RT-qPCR和流式细胞术评估了INS-1细胞的凋亡水平。Annexin-V-APC/7AAD双染流式细胞术结果显示,与BMSCs CM处理组相比,BMSCs-STS CM处理组凋亡细胞比例显著降低(图4B-C)。此外,BMSCs-STS CM预处理显著增加了Bcl-2蛋白表达,降低了Bax蛋白表达(图4D-E)。这些结果证实BMSCs-STS CM在体外保护INS-1细胞免于凋亡。
接下来,我们用两种CM处理胰岛24小时并收集细胞样本。与INS-1细胞中的结果一致,BMSCs-STS CM预处理显著增加了Bcl-2蛋白表达,降低了Bax蛋白表达(图4F-G)。我们还进行了AO/PI染色,观察到两种CM均降低了死细胞比例。然而,BMSCs-STS CM显著降低了PI染色强度,表明对胰岛具有更好的保护作用(图4H-I)。总之,BMSCs-STS CM预处理可在体外减少β细胞凋亡。
5.蛋白质组学揭示BMSCs-STS CM中的有益蛋白因子
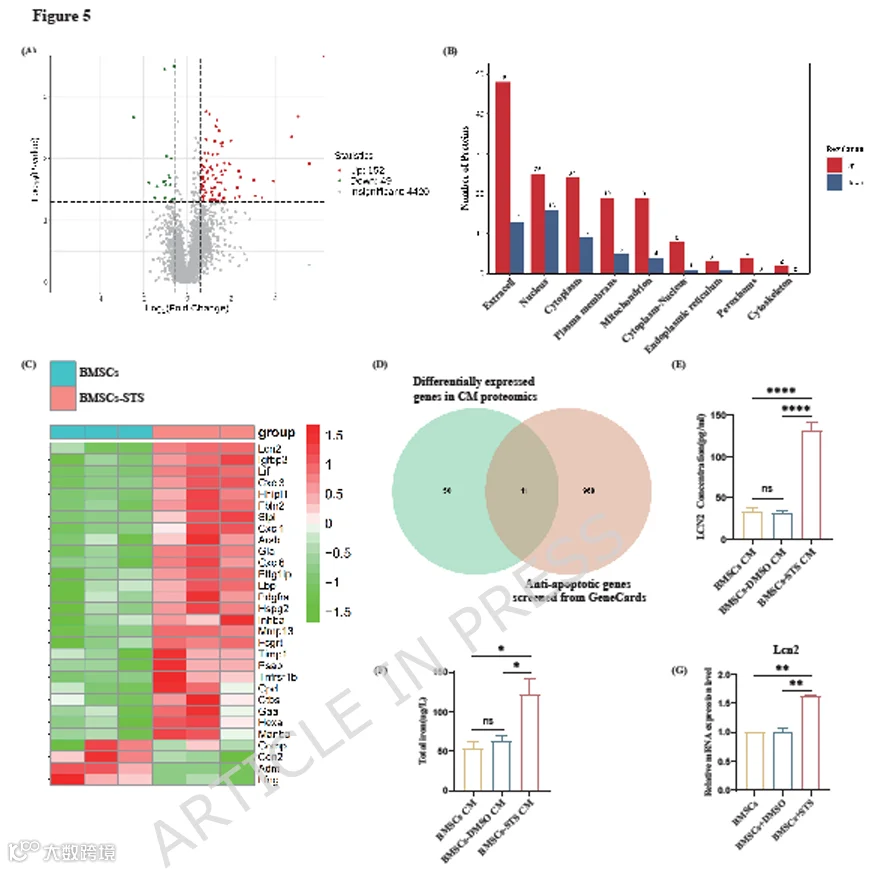
为了探索凋亡BMSCs在体内外减少β细胞凋亡的潜在机制,我们对BMSCs-STS CM和BMSCs CM进行了蛋白质组学测序(n=3)。结果显示,两组CM之间共有201个差异表达蛋白(DEPs)(列表见表S3),其中152个蛋白上调,49个蛋白下调(图5A)。由于MSCs主要通过旁分泌机制发挥其生物学效应,我们进一步分析了201个DEPs,以特别关注定位于细胞外或鉴定为分泌蛋白的蛋白,阈值为倍数变化≥1.5且p≤0.05(图5B-C)。然后我们在GeneCards数据库(https://www.genecards.org)中搜索抗凋亡蛋白(列表见表S4),并将其与DEPs中的分泌蛋白取交集,最终得到11个蛋白(图5D)。这些蛋白是:Slpi、Lcn2、Tnfsf1b、Timp1、IL6、Ngf、Adm、CCL20、Osm、IL11和Vcan。
其中,Lcn2的差异倍数最大。我们用1 ng/mL、10 ng/mL、100 ng/mL和500 ng/mL的重组Lcn2蛋白预处理INS-1细胞,然后用3 mM STZ诱导。通过CCK-8实验评估INS-1细胞活性。不幸的是,我们的实验结果表明,重组Lcn2蛋白预处理并未减轻INS-1细胞凋亡或增强其活性(图S2A)。此外,通过文献回顾,我们确定了其他具有高差异倍数的分泌蛋白,已知它们能保护细胞免于凋亡,如Lif和Psp。然而,用它们的重组蛋白预处理并未对INS-1细胞表现出保护作用(图S2B-C)。
有趣的是,我们发现先前的研究报道,铁负载型Lcn2(holo-Lcn2)可以在HeLa细胞、原代小鼠肾小管上皮细胞和肾肿瘤管状上皮细胞中保护细胞免受凋亡和铁死亡。然而,holo-Lcn2是否能抑制β细胞凋亡尚不清楚。
6.Holo-Lcn2预处理可抑制β细胞凋亡和铁死亡
为了验证holo-Lcn2是否为BMSCs-STS CM中有效的抗凋亡成分,我们首先使用ELISA检测了两种CM中Lcn2蛋白水平,观察到BMSCs-STS CM中Lcn2蛋白含量显著增加(图5E)。接下来,我们使用RT-qPCR证实,用0.2 μM STS处理的BMSCs中Lcn2的mRNA表达高于未处理的BMSCs(图5G),表明STS处理促进了BMSCs中Lcn2的分泌。此外,我们使用电感耦合等离子体质谱法测量了两种CM中的总铁水平,发现BMSCs-STS CM中的总铁水平显著高于BMSCs CM(图5F)。
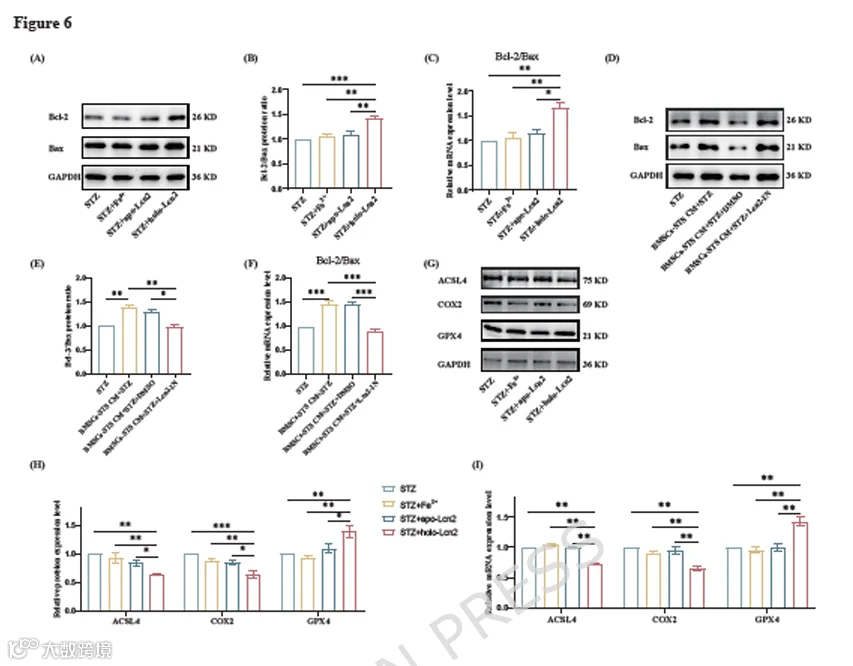
接下来,我们使用大肠杆菌菌株BL21(DE3)表达并纯化holo-Lcn2。然后用5 ng/mL的apo-Lcn2、holo-Lcn2或Fe³⁺(相同摩尔浓度)处理INS-1细胞24小时,再用3 mM STZ处理。通过Western blot和RT-qPCR评估INS-1细胞的凋亡水平。我们观察到holo-Lcn2增加了INS-1细胞中Bcl-2/Bax的蛋白和mRNA比率,从而抑制了细胞凋亡。然而,Fe³⁺和apo-Lcn2处理并未导致凋亡水平降低(图6A-C)。这些结果表明,只有铁负载型的Lcn2(holo-Lcn2)才能抑制INS-1细胞的凋亡。
此外,我们使用了Lcn2蛋白抑制剂ZINC00784494,该抑制剂通过直接结合Lcn2蛋白来拮抗Lcn2。将抑制剂加入BMSCs-STS CM后预处理INS-1细胞,我们再次通过Western blot和RT-qPCR评估凋亡。我们观察到加入ZINC00784494后,CM的抗凋亡效应被拮抗(图6D-F)。
考虑到添加的是铁负载型蛋白,且上述结果证明holo-Lcn2减轻了INS-1细胞凋亡,这引发了一个问题:它是否可能诱导其他形式的细胞死亡,如铁死亡。为此,我们通过Western blot和RT-qPCR检测了铁死亡相关标志物,发现用5 ng/mL holo-Lcn2处理并未诱导铁死亡,反而减轻了细胞中的铁死亡水平(图6G-I)。
总之,我们已经证明holo-Lcn2是BMSCs-STS CM中的一个关键抗凋亡因子。用holo-Lcn2预处理可以抑制β细胞凋亡并减轻铁死亡。
7.Holo-Lcn2通过Slc22a17将Fe³⁺转运入细胞内来抑制β细胞凋亡
为了确定holo-Lcn2如何抑制INS-1细胞凋亡,我们使用STRING数据库预测了与Lcn2相互作用的潜在蛋白。我们发现Slc22a17作为Lcn2的膜受体,可以与holo-Lcn2结合,将其内化进入细胞,同时将Fe³⁺转运入细胞内,从而抑制凋亡。基于此,我们进一步预测了潜在的机制,并在示意图中进行了说明(图Graphical Abstract),总之,holo-Lcn2通过将Fe³⁺转运入细胞内增加细胞内Fe³⁺水平来抑制凋亡。
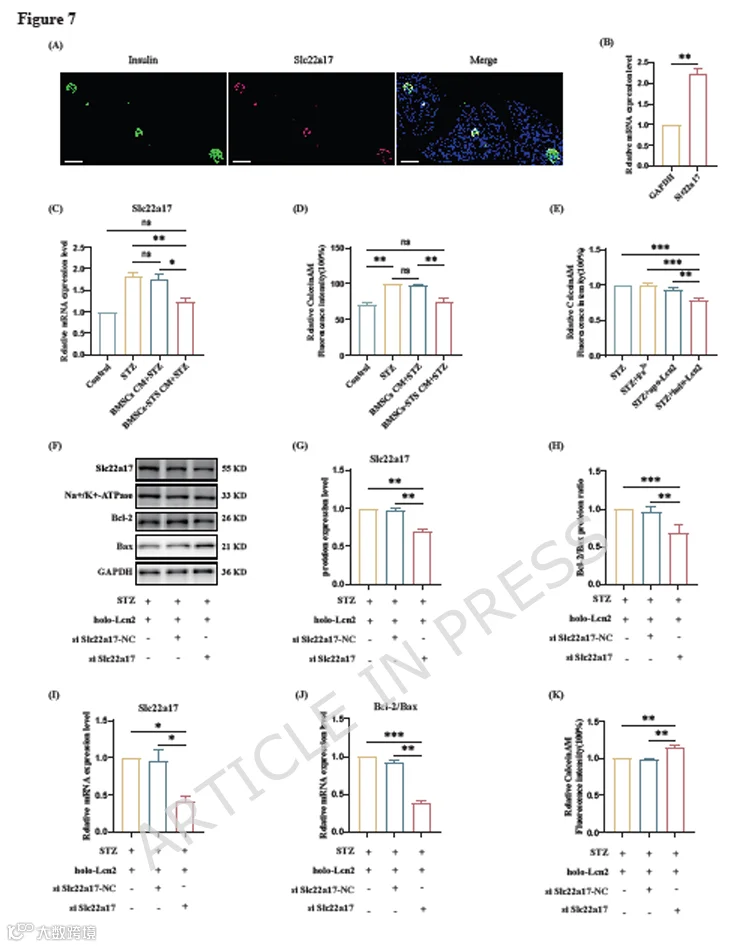
此前没有文献报道Slc22a17在胰岛或INS-1细胞中的表达。因此,我们首先提取大鼠胰腺组织并进行免疫荧光染色,发现分泌胰岛素的胰腺β细胞也表达Slc22a17(图7A)。然后我们分离胰岛,并在胰岛和INS-1细胞中进行RT-qPCR实验,以验证Slc22a17相对于GAPDH表达的mRNA水平(图7B-C)。这些实验证实了β细胞中存在Slc22a17表达。
接下来,我们使用了Fe³⁺染料Calcein-AM,它与Fe³⁺结合会导致荧光淬灭。我们对用两种CM、holo-Lcn2、apo-Lcn2和Fe³⁺预处理的INS-1细胞进行染色,发现用BMSCs-STS CM和holo-Lcn2处理的细胞内Fe³⁺含量增加(图7D-E)。
为了进一步研究Fe³⁺转运与凋亡缓解之间的相关性,我们进行了挽救实验。我们设计了三个Slc22a17-siRNA,其中siSlc22a17-2的敲低效率最显著(图S2D)。我们用siSlc22a17-2转染INS-1细胞,敲低Slc22a17,然后用holo-Lcn2预处理。随后评估细胞凋亡水平和Slc22a17表达。我们的研究结果显示,siSlc22a17组Bcl-2/Bax蛋白和mRNA比率没有增加,未能逆转INS-1细胞的凋亡(图7F-J),同时细胞内Fe³⁺水平也降低(图7K)。
总之,BMSCs-STS与胰岛共移植增加了移植物存活并减少了移植物凋亡。而且,我们证明只有铁负载形式的Lcn2才能与转运蛋白Slc22a17结合,然后将Fe³⁺转运入细胞内,从而减少β细胞凋亡。
更多结果和补充图表:doi: 10.1186/s13287-026-05001-9
长按二维码关注我们,用最短的时间和最高的效率学习更多生信思路!
扫描上方二维码或登录平台官网后添加CNSknowall客服微信咨询!官网地址:https://cnsknowall.com
CNSknowall:24年最新问世的遥遥领先的颠覆性科研数据(0代码生信+统计学)分析平台,同时含有机制图模块(原创3000多素材和机制图模板)+AI一键生成高质量比国自然标书初稿+汉化版Pubmed融合Deepseek高效筛选目标文献同时一键提炼全文核心创新点+SCI文献例句/语料检索模块+全文翻译+文献求助+图片查重+期刊查询+OPenAI官方GPT接口,>500款CNS级别图表皆可一秒内一键出图,登录即秒变数据分析大神,体验前所未有的便捷数据分析之旅,开启科研天骄之路!
可向下滑动发掘更多科研秘籍!